


Abstract
Aberrant ligand-independent G protein–coupled receptor constitutive activity has been implicated in the pathophysiology of a number of cancers. The adenosine A2B receptor (A2BAR) is dynamically upregulated under pathologic conditions associated with a hypoxic microenvironment, including solid tumors. This, in turn, may amplify ligand-independent A2BAR signal transduction. The contribution of A2BAR constitutive activity to disease progression is currently unknown yet of fundamental importance, as the preferred therapeutic modality for drugs designed to reduce A2BAR constitutive activity would be inverse agonism as opposed to neutral antagonism. The current study investigated A2BAR constitutive activity in a heterologous expression system and a native 22Rv1 human prostate cancer cell line exposed to hypoxic conditions (2% O2). The A2BAR inverse agonists, ZM241385 [4-(2-[7-amino-2-(2-furyl)[1,2,4]triazolo[2,3-a][1,3,5]triazin-5-ylamino]ethyl)phenol] or PSB-603 (8-(4-(4-(4-chlorophenyl)piperazide-1-sulfonyl)phenyl)-1-propylxanthine), mediated a concentration-dependent decrease in baseline cAMP levels in both cellular systems. Proliferation of multiple prostate cancer cell lines was also attenuated in the presence of PSB-603. Importantly, both the decrease in baseline cAMP accumulation and the reduction of proliferation were not influenced by the addition of adenosine deaminase, demonstrating that these effects are not dependent on stimulation of A2BARs by the endogenous agonist adenosine. Our study is the first to reveal that wild-type human A2BARs have high constitutive activity in both model and native cells. Furthermore, our findings demonstrate that this ligand-independent A2BAR constitutive activity is sufficient to promote prostate cancer cell proliferation in vitro. More broadly, A2BAR constitutive activity may have wider, currently unappreciated implications in pathologic conditions associated with a hypoxic microenvironment.
Introduction
Ligand-independent activation of G protein–coupled receptors (GPCRs), known as constitutive activity, is an established biologic phenomenon that results from the spontaneous isomerization of receptors from inactive to active states (Lefkowitz et al., 1993; Parra and Bond, 2007). The two-state model of receptor activation (Leff, 1995), which condenses the vast array of possible receptor conformations into either an active (R*) or inactive (R) form, provides the simplest conceptual framework for understanding constitutive activity and, in turn, inverse agonism. Inverse agonists selectively stabilize the R state of the receptor and thus inhibit both ligand-dependent and ligand-independent signal transduction. By contrast, agonists preferentially stabilize the R* state of the receptor, whereas neutral antagonists have equal affinity for both receptor states and only inhibit ligand-dependent effects (Strange, 2002; Milligan, 2003). Over the past three decades, the experimental capability to detect constitutive activity has enabled the reclassification of many clinically used antagonists as inverse agonists (Bond and Ijzerman, 2006). Although many GPCRs can be engineered or overexpressed to display some level of constitutive activity (Chalmers and Behan, 2002), the realization that mutant GPCRs with aberrant intrinsic activity can have a critical role in disease progression revealed the therapeutic potential for inverse agonists (Seifert and Wenzel-Seifert, 2002). Indeed, the degree of potential inverse agonism should be considered in the rational design and screening of drug candidates for disease states in which constitutive activity is known to be important (Milligan and IJzerman, 2000; Bond and IJzerman, 2006). As constitutive activity is linked to the proportion of receptors in the active receptor state (Parra and Bond, 2007), previously unidentified constitutive activity may also become evident in settings where receptor overexpression occurs as a direct consequence of disease pathology. This is particularly relevant for cancerous cells within solid tumors, where the hypoxic microenvironment promotes the upregulation of many receptors and proteins under the influence of hypoxia inducible factor 1-α (HIF-1α) control (Semenza, 2000; Subarsky and Hill, 2003).
The adenosine A2B receptor (A2BAR) is one of four structurally similar adenosine GPCRs that display distinct pharmacological profiles via differential coupling to Gi/o (A1AR and A3AR) or Gs (A2AAR and A2BAR) proteins. The A2BAR is a pleiotropically coupled GPCR, signaling via both Gs and Gq proteins (Linden et al., 1999; Fredholm et al., 2001a), and represents a key example of a highly dynamic GPCR whose expression is modified by disease. The A2BAR is significantly upregulated by HIF-1α in a number of cancers (Li et al., 2005), including human prostate cancer (Wei et al., 2013; Mousavi et al., 2015). Until recently, the A2BAR was presumed to have minor physiologic significance, in part due to its relatively low affinity for the endogenous agonist adenosine (Feoktistov and Biaggioni, 1997; Fredholm et al., 2001b). However the substantial increase in extracellular adenosine concentration (Sommerschild and Kirkebøen, 2000) and the upregulation of A2BAR expression under pathologic conditions such as hypoxia (Kong et al., 2006) and inflammation (Ham and Rees, 2008) suggest a possible maladaptive role of the A2BAR. Consequently, A2BAR antagonists are currently being explored as a novel therapeutic strategy for the treatment of inflammation (Ham and Rees, 2008), asthma and chronic obstructive pulmonary disease (Polosa and Blackburn, 2009), and diabetic nephropathy (Cárdenas et al., 2013). Importantly, recent studies have revealed a pivotal role for A2BAR signaling in cancer cell proliferation and progression of solid tumors of the bladder, breast, colon, and prostate (Ma et al., 2010; Cekic et al., 2012; Wei et al., 2013). However, no study to date has determined whether the dynamic regulation of the A2BAR results in constitutive activity, and whether this process, in addition to or independently of the influence of adenosine tone, contributes to any observed pathophysiology.
Thus, the current study profiled A2BAR constitutive activity within both a heterologous system and a native 22Rv1 human prostate cancer cell line, probing elevated basal activity with inverse agonists. To investigate the potential significance of constitutive activity on cancer pathophysiology, we subsequently examined the effect of the A2BAR inverse agonist 8-(4-(4-(4-chlorophenyl)piperazide-1-sulfonyl)phenyl)-1-propylxanthine (PSB-603) on the proliferation of two different prostate cancer cell lines in an environment depleted of extracellular endogenous adenosine. Our findings suggest that not only is the A2BAR constitutively active, but this ligand-independent activity is sufficient to drive prostate cancer cell proliferation. Thus, we have identified a novel mechanism by which the A2BAR contributes to disease pathology and supports the development of inverse agonists, rather than neutral antagonists, as potential A2BAR therapeutics.
Materials and Methods
Materials.
The AlphaScreen cAMP and SureFire phosphorylated extracellular signal-regulated kinase 1 and 2 (pERK1/2) kits, Ultima Gold scintillation cocktail, and cyclopentyl-1, 3-dipropylxanthine, 8-[dipropyl-2, 3-3H(N)] ([3H]DPCPX) were obtained from PerkinElmer Life and Analytical Sciences (Waltham, MA). The IP-One homogeneous time-resolved fluorescence (HTRF) kit was obtained from Cisbio Bioassays (Codolet, France). 22Rv1 and DU145 cells were purchased from American Type Culture Collection (ATCC; Manassa, VA). Hygromycin B and adenosine deaminase (ADA), derived from calf intestine, were obtained from Roche Diagnostics (Mannheim, Germany). Dulbecco’s modified Eagle’s medium (DMEM), fetal bovine serum (FBS), RPMI 1640 (ATCC 30-2001 modification) medium, high-capacity cDNA transcription kit, Annexin V–Alexa Fluor 488, penicillin/streptomycin, and trypsin were purchased from Life Technologies (Carlsbad, CA). The TaqMan gene expression assay kit was obtained from Applied Biosystems (Carlsbad, CA). Adenosine receptor antagonists PSB-603, 2-(2-furanyl)-7-[3-(4-methoxyphenyl)propyl]-7H-pyrazolo[4,3-e][1,2,4]triazolo[1,5-c]pyrimidin-5-amine (SCH 442416), and 4-(2-[7-amino-2-(2-furyl)[1,2,4]triazolo[2,3-a][1,3,5]triazin-5-ylamino]ethyl)phenol (ZM241385) were all purchased from Tocris Biosciences (Bristol, UK). The RNeasy plus mini kit was obtained from Qiagen (Valencia, CA), and the Pierce BCA protein assay kit, from Thermo Scientific (Rockford, IL). All other reagents were purchased from Sigma-Aldrich (St. Louis, MO) and were of analytical quality.
Cell Culture and Membrane Preparation.
FlpIn Chinese hamster ovary (FlpInCHO) cells, stably transfected with either the human A1, A2A, A2B, or A3 adenosine receptors (A1AR-FlpInCHO, A2AAR-FlpInCHO, A2BAR-FlpInCHO and A3AR-FlpInCHO, respectively) were grown in DMEM supplemented with 10% FBS and hygromycin B (500 μg/ml). The 22Rv1 human-derived prostate cancer cell line was maintained in RPMI 1640–ATCC 30-2001 medium containing 2 mM l-glutamine, 10 mM HEPES, 1 mM sodium pyruvate, 25 mM glucose, and 18 mM sodium bicarbonate supplemented with 10% FBS. The DU145 human-derived prostate cancer cell line was grown in RPMI medium supplemented with 10% FBS. All cells were maintained at 37°C in a 5% CO2 humidified incubator, grown to confluence, and then seeded into 96-well culture plates at assay-specific densities. For membrane preparation, A2BAR-FlpInCHO cells were grown to 90% confluence before being harvested with detaching buffer (10 mM HEPES, 7 mM EDTA, 150 mM NaCl, pH 7.4) and centrifuged (250 × g, 5 minutes). The cell pellet was resuspended in HEPES homogenization buffer (10 mM HEPES and 10 mM EDTA, pH 7.4) and homogenized using a hand-held homogenizer (Polytron; Kinematica, Littau-Lucerne, Switzerland) for four 5-second intervals interspersed with 30-second cooling on ice. The homogenate was centrifuged (40,000 × g, 30 minutes, 4°C). The cell pellet was resuspended in HEPES homogenization buffer, and homogenization and centrifugation were repeated. The cell pellet was then resuspended in HEPES assay buffer (10 mM HEPES, 0.1 mM EDTA, pH 7.4), and the protein content was determined using a Pierce BCA protein assay kit according to the manufacturer’s instructions.
Radioligand Binding.
Membrane homogenates of A2BAR-FlpInCHO cells (100 μg) were incubated in a 500-μl total volume of HEPES-buffered saline solution (25 mM HEPES, 10 mM glucose, 146 mM NaCl, 5 mM KCl, 1 mM MgSO4, 2 mM sodium pyruvate, and 1.3 mM CaCl2, pH 7.4) and 1 U/ml ADA at 37°C for 60 minutes. Homologous competition binding at the A2BAR-FlpInCHO was achieved by incubating membranes with [3H]DPCPX (3 or 10 nM) in the absence or presence of 0.3 nM to 10 μM 8-cyclopentyl-1,3-dipropylxanthine (DPCPX). Nonspecific binding was determined using 100 μM 5′-N-ethylcarboxamidoadenosine (NECA). Incubation was terminated by rapid filtration through 0.5% polyethylenimine presoaked Whatman GF/B filters using a membrane harvester (Brandel, Gaithersburg, MD). Filters were washed four times with 2 ml of ice-cold 0.9% NaCl, dried before the addition of 4 ml of scintillation cocktail (Ultima Gold; PerkinElmer Life and Analytical Sciences), and allowed to stand for 1 hour before radioactivity was determined by scintillation counting.
cAMP Accumulation.
FlpInCHO cells and 22Rv1 cells were seeded into 96-well plates at a density of 20,000 or 40,000 cells/well, respectively, and incubated in complete cell medium for 6 hours at 37°C in a humidified incubator. 22Rv1 cells were then placed in a sealed hypoxic chamber (Hypoxia Subchamber; BioSpherix, Lacona, NY) containing 2% O2 and 5% CO2 and incubated at 37°C for 24 hours. FlpInCHO and 22RV1 cell medium was removed and replaced with 200 μl/well culture medium containing 100 U/ml penicillin and 100 μg/ml streptomycin in the absence or presence of inverse agonist (0.1 nM to 10 μM) and/or 1 U/ml ADA, and cells were incubated for 16 hours at 37°C in a 5% CO2 humidified incubator. Medium was then removed, and cells were incubated with stimulation buffer (140 mM NaCl, 5 mM KCl, 0.8 mM MgSO4, 1.3 mM CaCl2, 0.2 mM Na2HPO4, 0.44 mM KH2PO4, 5.6 mM d-glucose, 1 mg/ml bovine serum albumin, 10 μM rolipram, and 5 mM HEPES, pH 7.4) in the absence and presence of 1 U/ml ADA and/or adenosine receptor ligand (concentrations ranging from 10 pM to 10 μM) for 30 minutes at 37°C. Stimulation was terminated by the removal of medium and the addition of 50 μl/well ice-cold ethanol. Detection of cAMP was performed using the AlphaScreen cAMP kit as described previously (Koole et al., 2010). Data were analyzed against a cAMP standard curve performed in parallel and expressed as cAMP concentration per well as a fold over basal.
Phosphorylation of Extracellular Signal-Regulated Kinase 1 and 2.
A2BAR-FlpInCHO or 22Rv1 cells were seeded into 96-well plates (40,000 cells/well) and allowed to adhere for 6 hours. 22Rv1 cells were placed in a sealed hypoxic chamber containing 2% O2 and 5% CO2 and incubated at 37°C for 24 hours. Cells were washed with phosphate-buffered saline and maintained in serum-free DMEM for 16 hours in the absence or presence of inverse agonists (concentrations ranging from 0.1 nM to 10 μM). Cells were then exposed to serum-free medium in the absence or presence of agonist for 5 minutes (concentrations ranging from 10 pM to 100 μM) followed by the removal of medium and addition of 100 μl/well SureFire lysis buffer to each well. Detection of pERK1/2 was performed using the AlphaScreen pERK1/2 SureFire kit as described previously (May et al., 2007). Data were normalized to the response elicited upon the exposure of cells to 10% FBS for 5 minutes.
Inositol Monophosphate Accumulation.
A2BAR-FlpInCHO cells in phenol red–free medium were seeded into sterilized 384-well ProxiPlates (PerkinElmer Life and Analytical Sciences) (12,000 cells/10 μl) and incubated for 16 hours in the absence or presence of inverse agonists. 22Rv1 cells were seeded into 96-well plates (20,000 cells/well) and placed in a sealed hypoxic chamber containing 2% O2 and 5% CO2 and incubated at 37°C for 24 hours. Where appropriate, cells were exposed to agonist for 1 hour prior to inositol monophosphate (IP1) being measured using the IP-One HTRF accumulation kit (Cisbio Bioassays), according to the manufacturer’s instructions, and detected with an EnVision microplate reader (PerkinElmer Life and Analytical Sciences) using standard HTRF settings (665-/630-nm ratio). Results were analyzed as an inverse ratio, with IP1 concentrations extrapolated from the IP1 standard curve performed in parallel.
Expression of Adenosine Receptor mRNA.
22Rv1 cells were harvested using 0.05% trypsin/0.53 mM EDTA solution. RNA was extracted using the RNeasy Plus Mini kit (Qiagen) according to the manufacturer’s instructions. RNA concentration was measured at 260 nm with a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE). Total RNA (2 µg) was then used for cDNA synthesis using the High Capacity cDNA Transcription Kit (LifeTechnologies, Carlsbad, CA) according to the manufacturer’s instructions. Reverse-transcription polymerase chain reaction (RT-PCR) validation was performed using TaqMan Gene Expression assays to detect and quantitate gene transcripts of human A1AR (TaqMan probe: Hs00379752_m1), A2AAR (TaqMan probe: Hs00169123_m1), A2BAR (TaqMan probe: Hs00386497_m1), and A3AR (TaqMan probe: Hs00252933_m1). In brief, cDNA samples were diluted 1:4 in nuclease-free water and mixed with TaqMan Fast Advanced Master Mix (Applied Biosystems). RT-PCR was performed using the Mastercycler ep realplex system (Eppendorf, Hamburg, Germany). Samples were analyzed in duplicate. The fluorescence threshold values were obtained, and calculation of relative change in mRNA was performed using the comparative delta delta cycle threshold method as described previously (Livak and Schmittgen, 2001) with normalization for the endogenous control β-actin (TaqMan probe: Hs01060665_g1).
Cell Proliferation and Cell Viability.
22Rv1 and DU145 cells were seeded in 96-well plates at 3000 cells/well in 200 μl of complete medium and cultured for 16 hours at 37°C. Cells were then exposed to complete medium in the absence or presence of inverse agonists PSB-603 (1 μM) or SCH 442416 (100 nM; 22Rv1 cells only) and/or 1 U/ml ADA for 24 or 48 hours. Cells were maintained in a hypoxic chamber containing 2% O2 and 5% CO2 at 37°C for the duration of the experiment. Medium, inverse agonists, and ADA were refreshed every 24 hours. Cell proliferation was determined after labeling with 3-(4,5- dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT). Absorbance of the converted dye was determined by subtraction of the background signal at 690 nm from the measured absorbance at 570 nm on a FlexStationIII plate reader (Molecular Devices, Sunnyvale, CA). 22Rv1 cell viability was determined with annexin V and propidium iodide (PI) staining. 22Rv1 cells, grown in petri dishes and exposed to the same conditions described for cell proliferation assays, were harvested in annexin-binding buffer (10 mM HEPES, 140 mM NaCl, and 2.5 mM CaCl2, pH 7.4) and stained with Annexin V–Alexa Fluor 488 (5/100 μl) and PI (5 μg/ml) for 15 minutes at room temperature. Samples were immediately analyzed using the BD fluorescence-activated cell sorter (FACS) CantoII flow cytometer (BD Biosciences, San Jose, CA).
Data Analysis.
All data were analyzed using Prism 6.0 (GraphPad Software Inc., San Diego, CA). Statistical significance was defined as P < 0.05 as determined by one-way or two-way analysis of variance (ANOVA) with Tukey’s or Bonferroni’s multiple comparisons post-hoc analysis or t test, as indicated within the results.
Results
The A2BAR-FlpInCHO Cell Line Has Elevated Basal cAMP.
Parental and adenosine receptor FlpInCHO cell lines, assessed in parallel, demonstrated significant differences in baseline cAMP accumulation. Specifically, the A2BAR-FlpInCHO cell line had significantly higher baseline cAMP when compared with the parental, A1AR-, A2AAR-, and A3AR-FlpInCHO cell lines (Fig. 1; P < 0.01, one-way ANOVA Tukey’s multiple comparison test, n = 5). Receptor number can influence basal cAMP accumulation through constitutive activation of adenylyl cyclase (Nakahara et al., 2004); as such, we sought to determine the level of adenosine receptor expression in the stably transfected A2B-FlpInCHO cell line. [3H]DPCPX homologous competition binding on membrane homogenates of A2BAR-FlpInCHO cell membranes yielded an affinity estimate (pKd of 6.87 ± 0.10; Supplemental Fig. 1) that was similar to published values (Weyler et al., 2006). Importantly, the A2BAR was not grossly overexpressed, with a Bmax value of 3.13 ± 0.61 pmol mg protein−1, which is comparable to other GPCRs expressed in the FlpInCHO cell system (Yan et al., 2009; Gregory et al., 2010).

Human A2BAR-FlpInCHO cells have a higher level of baseline cAMP than the parental or other AR-FlpInCHO cells. Basal levels of cAMP accumulation in nontransfected (NT), A1AR-, A2AAR-, A2BAR-, and A3AR-FlpInCHO cells. **P < 0.01, one-way ANOVA; Tukey’s multiple comparison test. Data represent the mean + S.E.M. from five independent experiments performed in triplicate.
Concentration-Response Relationships for Adenosine Receptor Agonists and Inverse Agonists Reveal A2BAR Constitutive Activity for Gs-Coupled cAMP Accumulation.
In A2BAR-FlpInCHO cells, a robust and concentration-dependent stimulation of cAMP accumulation was observed for the nonselective adenosine receptor agonists, NECA and adenosine (Fig. 2A), with NECA 80-fold more potent than adenosine with respect to cAMP stimulation (Table 1). A2BAR constitutive activity was assessed by exposure of A2BAR-FlpInCHO cells to the A2BAR inverse agonists PSB-603 and ZM241385. PSB-603 and ZM241385 both mediated a significant and concentration-dependent reduction in baseline cAMP accumulation (Fig. 2A; P < 0.001, paired t test, n = 4–7). The observed decrease in baseline cAMP accumulation in the presence of inverse agonist is consistent with either inhibition of endogenous agonist activity or a reduction in constitutive activity. To investigate the influence of endogenous adenosine, concentration-response curves to agonists and inverse agonists were repeated in the presence of 1 U/ml ADA. ADA had no significant effect on the potency (pEC50) or maximal effect (Emax) of NECA, but abolished the cAMP accumulation mediated by exogenous adenosine up to 10 μM (Fig. 2B; Table 1; P > 0.05, unpaired t test, no statistical significance for pEC50 or Emax of NECA ± ADA, n = 4–7). The concentration-dependent decrease in cAMP accumulation observed upon exposure of A2BAR-FlpInCHO cells to PSB-603 or ZM241385 was maintained in the presence of ADA (Fig. 2B; P < 0.001, paired t test, n = 4–7). Furthermore, the potency and maximal effect of ZM241385 or PSB-603 were not significantly different from that observed in the absence of ADA (Table 1; P > 0.05, unpaired t test, n = 4–7).

Inverse agonism of cAMP accumulation reveals A2BAR constitutive activity. (A) In the absence of ADA, exposure of A2BAR-FlpInCHO cells to the prototypical adenosine receptor agonists NECA or adenosine mediates robust increases in cAMP, whereas the A2BAR inverse agonists ZM241385 or PSB-603 significantly decrease baseline cAMP levels. (B) In the presence of 1 U/ml ADA, NECA mediates a robust increase in cAMP accumulation, the A2BAR inverse agonists ZM241385 or PSB-603 significantly decrease baseline cAMP levels, whereas the response to adenosine is abolished. ***P < 0.001; paired t test. Data represent the mean + S.E.M. from four to seven independent experiments performed in triplicate. Error bars not shown lie within the dimensions of the symbol.
Potency (pEC50) and maximal response (Emax) of A2BAR-mediated changes in cAMP accumulation in the presence and absence of ADA in A2BAR-FlpInCHO cells
Data are the mean ± S.E.M. from four or more independent experiments performed in triplicate.
A2BAR Constitutive Activity Cannot Be Detected at Alternative Signal Transduction Pathways, ERK1/2 Phosphorylation, and IP1 Accumulation.
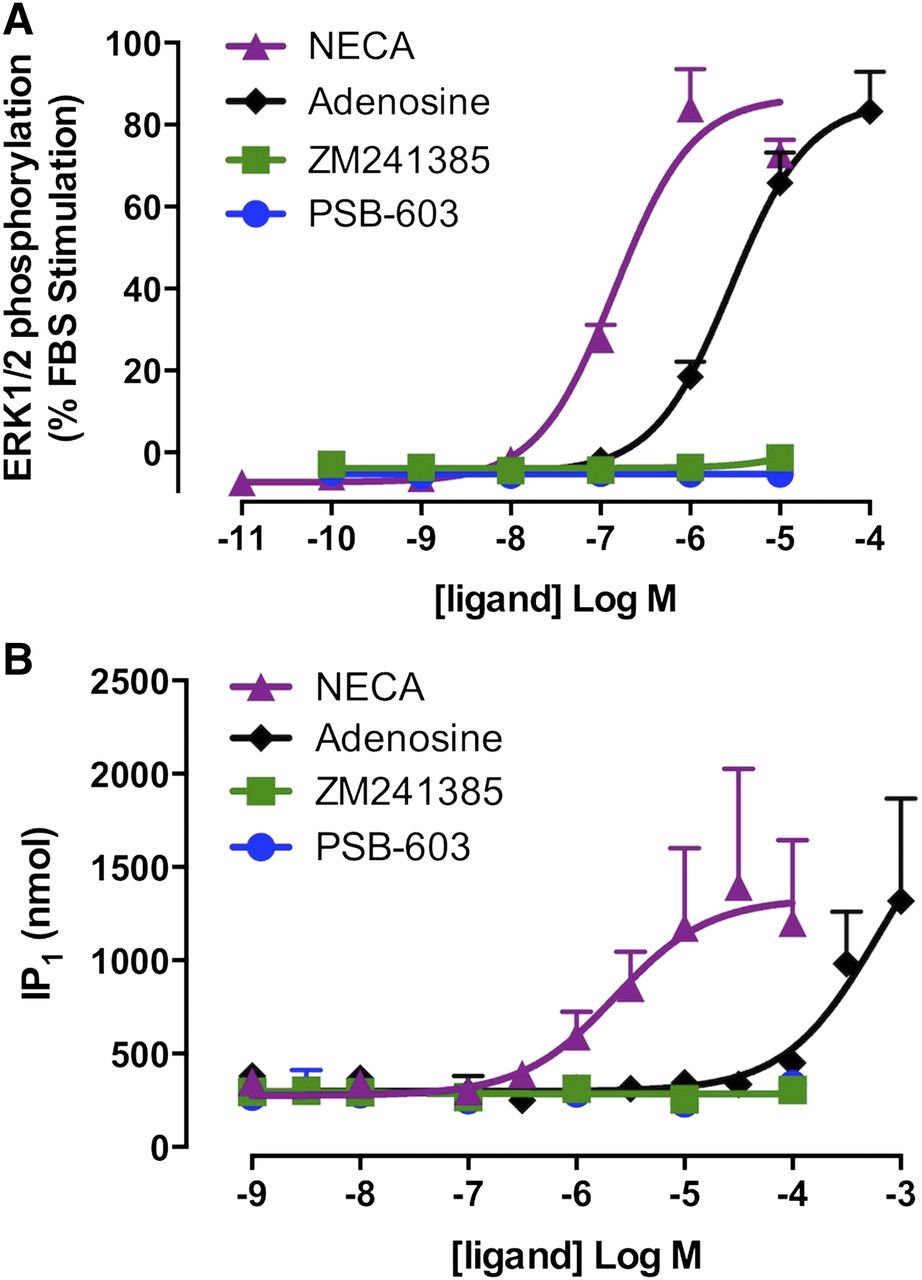
The A2BAR is a pleiotropically coupled GPCR, signaling via both Gs and Gq proteins (Linden et al., 1999; Fredholm et al., 2001a). To investigate the constitutive activation of alternate A2BAR signaling pathways, the influence of agonists and inverse agonists on baseline pERK1/2 (Fig. 3A) and IP1 accumulation (Fig. 3B) was assessed. Concentration-dependent increases in pERK1/2 and IP1 accumulation in the A2BAR-FlpInCHO cells were observed in the presence of NECA or adenosine, albeit at lower potencies than those observed in the cAMP accumulation assay (Fig. 3). Specifically, the potency (pEC50) of NECA-mediated pERK1/2 and IP1 accumulation was 6.69 ± 0.07 and 5.74 ± 0.20, respectively, whereas the pEC50 for adenosine-mediated pERK1/2 was 5.55 ± 0.07 and could not be determined for IP1 accumulation. The inverse agonists, PSB-603 or ZM241385, had no effect on baseline pERK1/2 or IP1 accumulation at any concentration assessed (0.1 nM to 100 μM) (Fig. 3). Concentration-dependent increases in pERK1/2 and IP1 accumulation in the presence of NECA and adenosine could not be detected in 22Rv1 cells (Supplemental Fig. 2).

Inverse agonism was not observed for A2BAR-mediated ERK1/2 phosphorylation (A) or IP1 accumulation (B) in A2BAR-FlpInCHO cells. Data represent the mean + S.E.M. from four independent experiments performed in triplicate. Error bars not shown lie within the dimensions of the symbol.
Hypoxia-Inducible A2BARs in the 22Rv1 Human Prostate Cancer Cell Line Are Constitutively Active.
RT-PCR analysis of the 22Rv1 prostate cancer cells detected a high level of A2AAR mRNA, a moderate level of A2BAR mRNA, and a low level of A1AR mRNA (Fig. 4A). A3AR mRNA was below the level of detection. As demonstrated previously in other human carcinoma cell lines (Kong et al., 2006; Ma et al., 2010), expression of A2BAR mRNA was significantly upregulated after 8 hours of hypoxia (2% O2/5% CO2; P < 0.01, two-way ANOVA Bonferroni’s multiple comparison test, n = 3). In contrast, no significant differences were observed in the level of A1AR and A2AAR mRNA detected under normoxic conditions or after 8 hours of hypoxia (Fig. 4B).
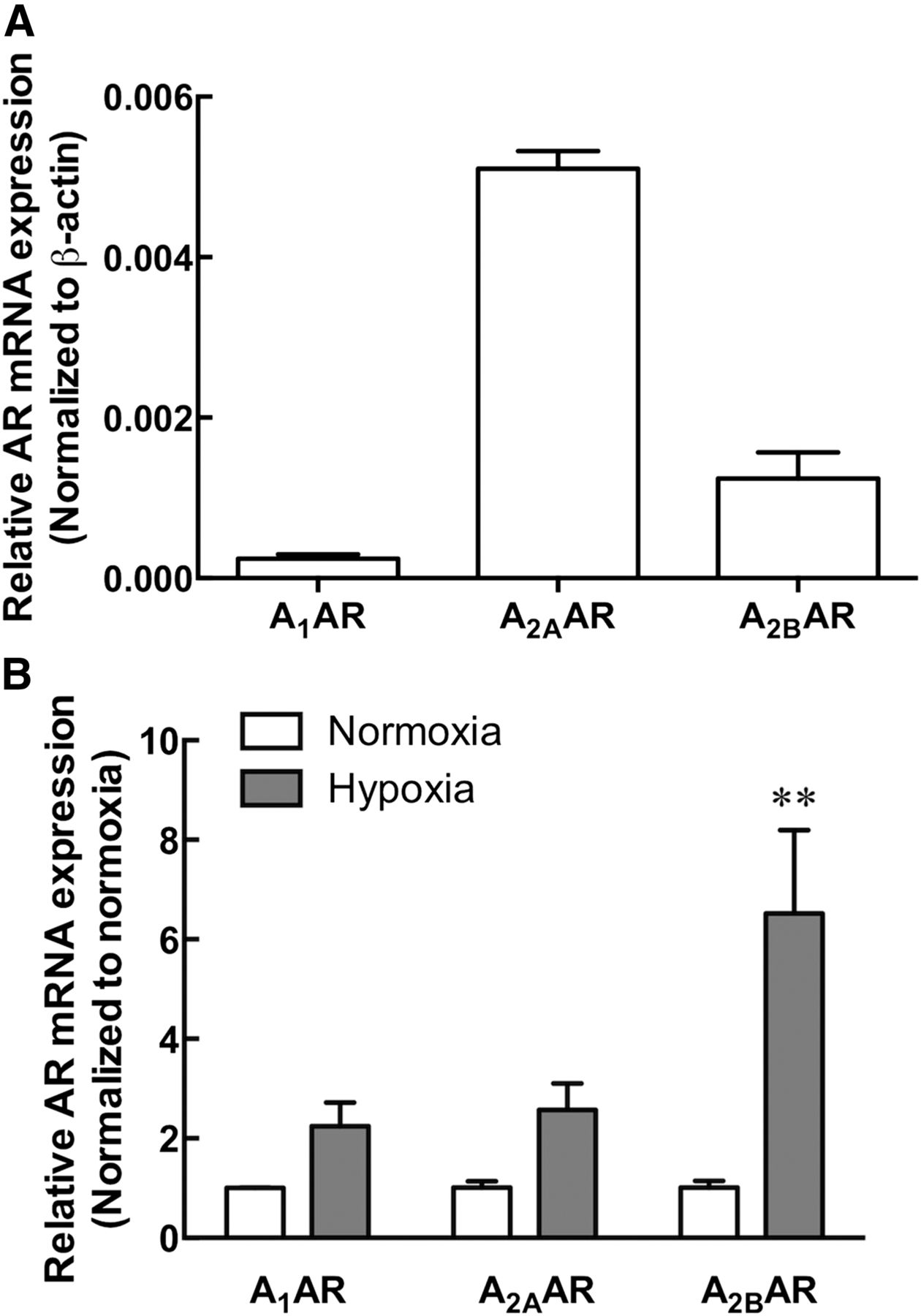
A2BAR mRNA expression is upregulated under hypoxic conditions in the 22Rv1 prostate cancer cell line. (A) The relative expression of human A1AR, A2AAR, and A2BAR mRNA in 22Rv1 prostate cancer cells under normoxic conditions. A3AR mRNA was below the level of detection. (B) A1AR, A2AAR, and A2BAR mRNA expression after exposure of 22Rv1 prostate cancer cells to 8-hour hypoxia (2% O2/5% CO2) normalized to respective normoxic control. **P < 0.01, two-way ANOVA, Bonferroni’s multiple comparison test. Data represent the mean + S.E.M. from three independent experiments.
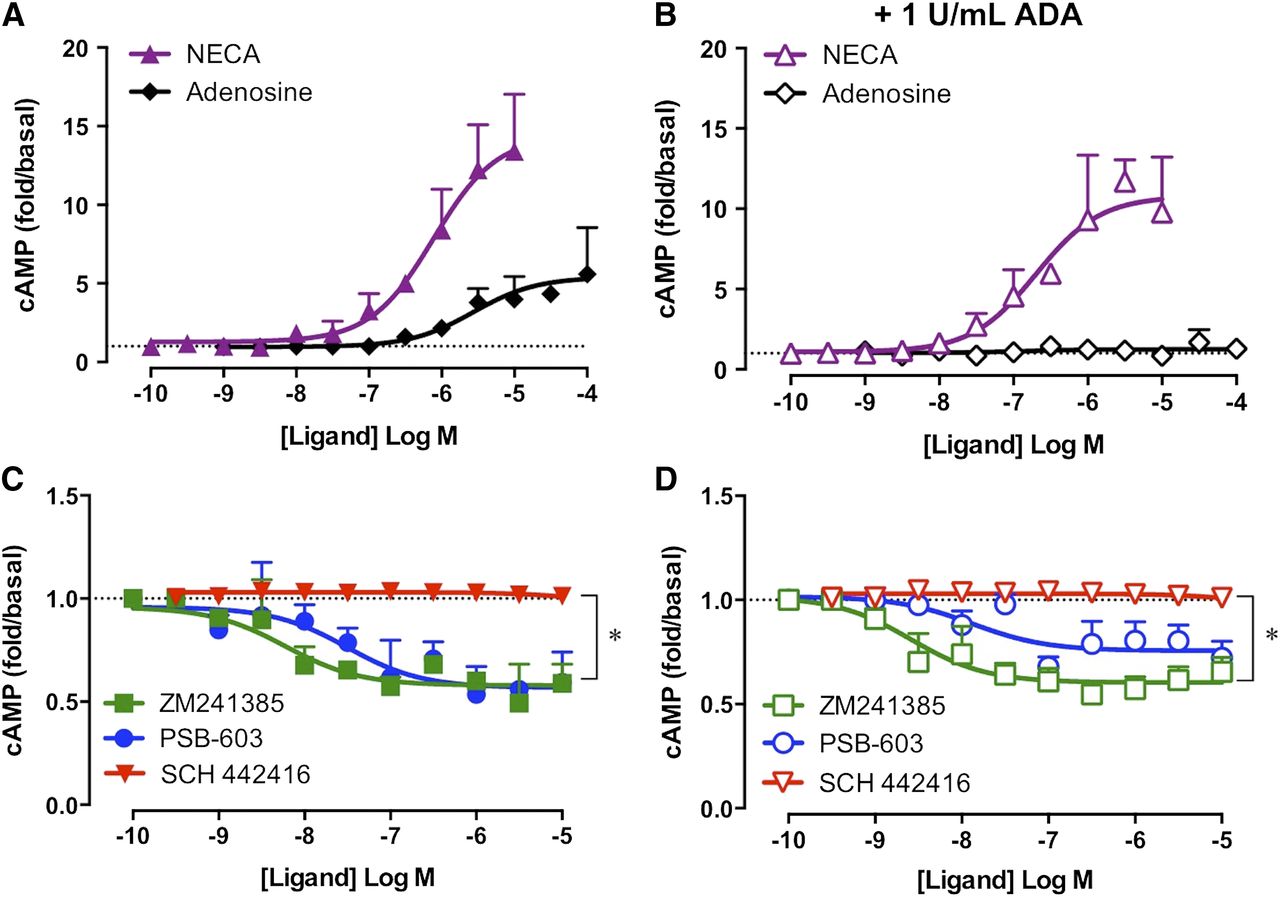
Constitutive A2BAR activity in 22Rv1 prostate cancer cells was investigated after exposure of cells to hypoxia, the condition for which increased A2BAR expression is likely to be observed. Specifically, the influence on cAMP accumulation in 22Rv1 prostate cancer cells was quantified for the nonselective adenosine receptor agonists NECA or adenosine, the A2BAR inverse agonists PSB-603 or ZM241385, and the A2AAR-selective antagonist SCH 442416 after 24 hours of hypoxia (2% O2/5% CO2). The A2AAR-selective antagonist, SCH 442416, was used to evaluate the influence the highly expressed A2AAR had on baseline cAMP. Robust concentration-dependent increases in cAMP were detected in both the absence and presence of 1 U/ml ADA for NECA and in the absence of ADA for adenosine (Fig. 5, A and B). The A2AAR-selective inverse agonist SCH 442416 had no effect on baseline cAMP under either condition tested (Fig. 5, C and D). In the absence of ADA, ZM241385 or PSB-603 mediated concentration-dependent decreases in basal cAMP with similar potencies (Fig. 5C). Furthermore, the small but significant window of inverse agonism was maintained in the presence of ADA (Fig. 5D; P < 0.05, paired t test, n = 3–4). The ability of ZM241385 or PSB-603 to inhibit ligand-independent cAMP accumulation in the 22Rv1 cells with similar potency to that observed in the A2BAR-FlpInCHO cells (ZM241385 pEC50: 8.44 ± 0.45 and 8.00 ± 0.26, respectively; PSB-603 pEC50: 7.67 ± 0.30 and 8.33 ± 0.38, respectively) reveals that A2BAR constitutive activity can be observed in both model heterologous expression systems and endogenous expression systems.
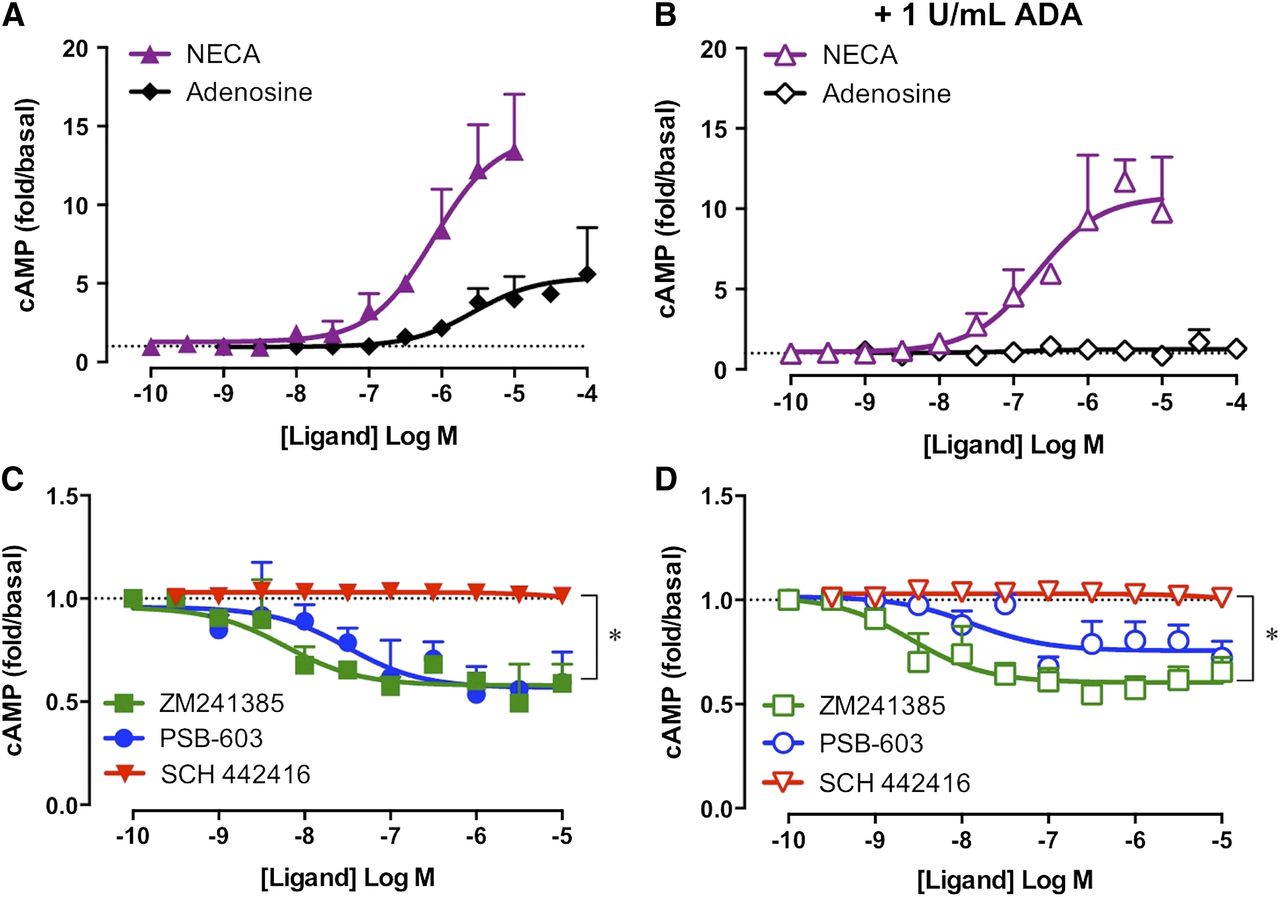
A2BARs endogenously expressed in 22Rv1 prostate cancer cells are constitutively active. (A) In the absence of ADA, the prototypical A2BAR agonists NECA or adenosine mediate robust increases in cAMP after 30-minute stimulation. (B) In the presence of ADA, NECA mediates a robust increase in cAMP accumulation, whereas the response to adenosine is abolished. (C and D) The A2AAR antagonist SCH 442416 had no effect on baseline cAMP accumulation. The A2BAR inverse agonists ZM241385 and PSB-603 mediate small but significant decreases in baseline cAMP in the absence (C) and presence (D) of 1 U/ml ADA. 22Rv1 cells were exposed to 24-hour hypoxia (2% O2/5% CO2) immediately prior to detection of cAMP accumulation. *P < 0.05, paired t test. Data represent the mean + S.E.M. from three to four independent experiments performed in triplicate. Error bars not shown lie within the dimensions of the symbol.
A2BAR Constitutive Activity Promotes Proliferation of 22Rv1 Human Prostate Cancer Cells.
The influence of A2BAR constitutive activity on the proliferation and survival of the 22Rv1 human prostate cancer cell line was investigated under hypoxic conditions (2% O2/5% CO2). As described previously, 1 U/ml ADA (refreshed every 24 hours) was used to investigate the influence of endogenous adenosine. Importantly, the activity of ADA after 24 hours at 37°C was confirmed in signal transduction assays (Supplemental Fig. 3). Over a 48-hour period, the A2BAR inverse agonist, PSB-603 (1 μM), significantly reduced 22Rv1 cell proliferation when compared with the respective buffer control, both in the absence and presence of ADA, determined using an MTT proliferation assay (Fig. 6A; P < 0.01, two-way ANOVA Tukey’s multiple comparison test, n = 5). Similar results were also observed in the DU145 prostate cancer cell line, whereby PSB-603 (1 μM) significantly reduced basal cell proliferation at 48 hours (Supplemental Fig. 4; P < 0.05, two-way ANOVA Tukey’s multiple comparison test, n = 4). A limitation of the MTT assay is that it does not differentiate between cell quiescence and increased cell death (Carmichael et al., 1987). Therefore, the 22Rv1 cells were also subjected to annexin V and PI staining and analyzed via FACS to assess whether PSB-603 had a significant influence on cell viability. The inverse agonist, PSB-603 (1 μM), had no significant effect on the percentage of nonviable annexin V or PI-positive cells after 48 hours (Fig. 6B; P > 0.05, two-way ANOVA Tukey’s multiple comparison test, n = 3). Thus, the reduced absorbance observed in the MTT assay is likely to be due to PSB-603 causing a decrease in proliferation as opposed to having a cytotoxic effect. The level of A2AAR mRNA was relatively high in 22Rv1 cells (Fig. 4A). To investigate whether inhibition of the A2AAR could also influence the proliferation of 22Rv1 cells, cells were exposed to the highly selective A2AAR antagonist, SCH 442416 (100 nM), and assessed in the MTT assay as described earlier. Over a 48-hour period, SCH 442416 (100 nM) had no significant effect on 22Rv1 cell proliferation (Fig. 6C; P > 0.05, two-way ANOVA Tukey’s multiple comparison test, n = 5).

The A2BAR inverse agonist PSB-603 decreased proliferation of 22Rv1 prostate cancer cells over 48 hours. (A) PSB-603 (1 μM) significantly decreased the proliferation of 22Rv1 cells in the absence (closed symbols) and presence (open symbols) of 1 U/ml ADA as determined by MTT absorbance. **P < 0.01 compared with respective buffer control; two-way ANOVA, Tukey’s multiple comparison test. (B) The A2BAR inverse agonist PSB-603 (1 μM) has no effect on cell viability after 48 hours, as assessed by the proportion of annexin V, PI, or double positive cells, in the absence and presence of 1 U/ml ADA. (C) The A2AAR-selective antagonist SCH 442416 (100 nM) had no effect on the proliferation of 22Rv1 cells in the absence (closed symbols) or presence (open symbols) of 1 U/ml ADA. 22Rv1 cells seeded at 3000 cells/well and maintained under hypoxia (2% O2/5% CO2), with medium, drugs, and ADA refreshed every 24 hours. Data represent the mean + S.E.M. from five (A and C) or three (B) independent experiments performed in triplicate. Error bars not shown lie within the dimensions of the symbol. +ve, positive.
Discussion
This study is the first to characterize the constitutive activity of the human wild-type A2BAR within both a heterologous FlpInCHO system and the native 22Rv1 prostate cancer cell line. Constitutive activity of the A2BAR was revealed through the detection of inverse agonism of cAMP accumulation under conditions that removed extracellular endogenous agonist. Furthermore, we identified a pathophysiological consequence for the elevated ligand-independent activity. That is, A2BAR constitutive activity stimulated proliferation of the 22Rv1 cell line, a finding that was confirmed in the DU145 prostate cancer cell line. As such, our studies indicate that future drug discovery efforts should differentiate A2BAR inverse agonists from neutral antagonists, as the former may prove a better potential therapeutic approach to slow prostate tumor progression. In addition, our studies suggest that ligand-independent constitutive activity should be taken into consideration when investigating the role of A2BARs in other disease pathologies.
Experimental observations for A2BAR ligand-mediated changes in cAMP accumulation are in accordance with a constitutively active system described by the two-state model of receptor activation (Leff, 1995; Bennett et al., 2013). In this model, the intrinsic efficacy of a ligand is governed by its relative affinity for the R verses R* state (Burstein et al., 1997). As such, an increase in agonist potency but a decrease in inverse agonist potency is predicted for a receptor system that has a high proportion of receptors existing in the R* state—that is, a constitutively active receptor system. In agreement with the two-state model, the NECA potency in the cAMP accumulation assay is approximately 100-fold higher than its affinity at the R state as estimated using antagonist 125I-3-(4-amino-3-iodobenzyl)-8-(phenyl-4-oxyacetate)-1-propylxanthine ([125I]ABOPX) binding (Linden et al., 1999). Conversely the functional potency of the inverse agonist PSB-603 is 10-fold lower than the R-state affinity as estimated using homologous [3H]PSB-603 competition binding (Borrmann et al., 2009). Therefore, the experimentally observed shift in agonist and inverse agonist potencies in the present study is in accordance with the two-state model and supports the suggestion that a significant proportion of A2BARs exist in the R* state.
Constitutive activity has previously been demonstrated for GPCRs coupling to each of the different heterotrimeric G proteins (Barker et al., 1994; Neilan et al., 1999; Hopkinson et al., 2000). Because A2BAR constitutive activity was only observed in the cAMP accumulation assay, this may reflect preferential ligand-independent stabilization of a Gs-coupled active receptor conformation. This contention is supported by studies of other pleiotropically coupled GPCRs, for which constitutive activity manifests exclusively in the Gs-coupled pathway, such as the gain–of-function mutation in the luteinizing hormone receptor responsible for precocious puberty and the hyperthyroidism-causing mutation of the thyroid-stimulating hormone receptor (Kopp et al., 1995; Liu et al., 1999; Seifert and Wenzel-Seifert, 2002). However, it is important to note that NECA and adenosine have a 20- to 2000-fold higher potency for cAMP when compared with IP1 and pERK1/2 in the A2BAR-FlpInCHO cell background and no detectable Gq-mediated activity in the 22Rv1 cells, suggesting that the A2BAR has a higher coupling efficiency for Gs- rather than Gq-coupled signaling. Therefore, it is possible that the cAMP accumulation assay is the only method sensitive enough to detect small changes below a baseline level (Seifert and Wenzel-Seifert, 2002). Regardless of mechanism, it is apparent that the elevated baseline cAMP is attributable to constitutive coupling to Gs proteins and supports the possibility for A2BAR constitutive activity pathway bias.
To determine whether the A2BAR constitutive activity observed in the heterologous system was relevant in an endogenous setting, we first used the 22Rv1, xenograft-derived human prostate carcinoma epithelial cell line. This cell line was selected based on previous reports of endogenous A2BAR expression within prostate cancer cells (Wei et al., 2013). Under hypoxic conditions, such as those observed within a tumor microenvironment (Subarsky and Hill, 2003; Ryzhov et al., 2008), A2BAR expression is increased by HIF-1α (Kong et al., 2006; Eckle et al., 2014). Adenosine receptor–mediated cAMP accumulation was assessed after 24 hours hypoxia (2% O2/5% CO2) to allow for A2BAR upregulation, thus enhancing the ability to detect constitutive activity (Nakahara et al., 2004). The nonselective agonists, NECA or adenosine, produced substantial increases in cAMP, likely due to the nonselective stimulation of A2BARs and A2AARs, both of which preferentially couple to Gs proteins. The A2BAR inverse agonists ZM241385 and PSB-603 produced a concentration-dependent decrease in baseline activity in the presence and absence of ADA, whereas the A2AAR-selective antagonist SCH 442416 had no effect on baseline cAMP levels. Given adenosine has a higher affinity for the A2AAR than the A2BAR (Liang and Haltiwanger, 1995; Fredholm et al., 2001b), we would anticipate that, if ZM241385 and PSB-603 were simply inhibiting the effects of residual endogenous adenosine, then a comparable or larger effect on baseline cAMP should be observed in the presence of an A2AAR-selective competitive antagonist. As such, the absence of an effect in the presence of A2AAR blockade argues against the possibility of a contaminating influence of endogenous adenosine and instead further supports appreciable A2BAR constitutive activity in the 22Rv1 prostate cancer cell line.
A2BAR expression at both the mRNA and protein level is higher in malignant prostate cancer tissue from human patients when compared with normal control prostate tissue (Mousavi et al., 2015). In addition, the A2BAR has been implicated in cell proliferation and angiogenesis, accounting for its apparent role in the pathogenesis of a number of solid tumors (Li et al., 2005; Ryzhov et al., 2008; Ma et al., 2010; Cekic et al., 2012; Iannone et al., 2013; Wei et al., 2013). The involvement of the A2BAR in the proliferation of human oral squamous cell carcinoma–derived cells was confirmed with small hairpin RNA (Kasama et al., 2015). Furthermore, previous studies have demonstrated the inhibitory effects of an A2BAR inverse agonist on the cell proliferation of human colon carcinomas (Ma et al., 2010) and prostate cancer cells (Wei et al., 2013); however, these studies were performed in the presence of endogenous adenosine and thus were unable to differentiate between the influence of agonist tone and any potential ligand-independent receptor activity. We sought to directly determine whether A2BAR constitutive activity contributed to the basal level of cell growth of prostate cancer cells. Our results revealed that, even when endogenous agonist had been removed, the A2BAR inverse agonist PSB-603 significantly suppressed cell growth of malignant 22Rv1 and DU145 prostate cells over the 48-hour period. The A2AAR-selective antagonist, SCH 442416, did not replicate the inhibition of cell proliferation in 22Rv1 cells, supporting the supposition that A2BAR constitutive activity, and not endogenous adenosine, significantly contributes to pathologic prostate cancer cell proliferation in vitro. Differentiating the effect of constitutive activity from the effect of the endogenous ligand in an in vivo setting is challenging (Parra and Bond, 2007). However, targeting pathologic A2BAR overexpression and concurrent constitutive activity with inverse agonists is readily achievable and presents an exciting future prospect in cancer treatment.
In conclusion, ligand-independent A2BAR constitutive activity of the Gs-coupled cAMP pathway can be detected in both a heterologous and native cell line. The effect of an A2BAR inverse agonist to reduce the basal level of cell growth of two different prostate cancer cell lines demonstrates the potential therapeutic benefit in targeting A2BAR constitutive activity as a pharmacological adjuvant in prostate cancer treatment. Furthermore, this study highlights the requirement to differentiate potential inverse agonist effects from neutral antagonist effects of A2BAR compounds in the drug discovery pipeline to identify optimal therapeutically efficacious ligands for this receptor. Although our study demonstrates the pathologic relevance of A2BAR constitutive activity within the context of prostate cancer, it may also have wider implications in both physiologic and pathologic conditions where the A2BAR was previously considered to have minimal influence.
Acknowledgments
The authors thank Dr. Michael Crouch and Dr. Ron Osmond for providing the pERK1/2 assay kit, Dr. Gail Risbridger for assistance with the DU145 prostate cancer cells, and Sandy Fung for assistance with FACS analysis.
Authorship Contributions
Participated in research design: Vecchio, White, May.
Conducted experiments: Vecchio, Tan.
Performed data analysis: Vecchio, White, May.
Wrote or contributed to the writing of the manuscript: Vecchio, Gregory, Christopoulos, White, May.
Footnotes
- Received October 13, 2015.
- Accepted January 19, 2016.
This work was supported by the National Health and Medical Research Council (NHMRC) of Australia [1084487] and the Australian Research Council [DE130100117]. L.T.M. is an Australian Research Council (ARC) Discovery Early Career Researcher Award (DECRA) Postdoctoral Research Fellow [DE130100117], K.J.G. is an NHMRC CJ Martin Fellow [1013709], and A.C. is an NHMRC Principal Research Fellow [1041875]. E.A.V. holds an Australian Postgraduate Award and an Australian Cancer Therapeutics scholarship.
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- A2BAR
- adenosine A2B receptor
- ADA
- adenosine deaminase
- ANOVA
- analysis of variance
- ATCC
- American Type Culture Collection
- DMEM
- Dulbecco’s modified Eagle’s medium
- DPCPX
- 8-cyclopentyl-1,3-dipropylxanthine
- Emax
- maximal effect
- FACS
- fluorescence-activated cell sorter
- FBS
- fetal bovine serum
- FlpInCHO
- Chinese hamster ovary cells with FlpIn vector
- GPCR
- G protein–coupled receptor
- [3H]DPCPX
- cyclopentyl-1, 3-dipropylxanthine, 8-[dipropyl-2, 3-3H(N)]
- HIF-1α
- hypoxia inducible factor 1-α
- HTRF
- homogeneous time-resolved fluorescence
- IP1
- inositol monophosphate
- MTT
- 3-(4,5- dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
- NECA
- 5′-N-ethylcarboxamidoadenosine
- pEC50
- potency
- pERK1/2
- phosphorylated extracellular signal-regulated kinase 1 and 2
- PI
- propidium iodide
- PSB-603
- 8-(4-(4-(4-chlorophenyl)piperazide-1-sulfonyl)phenyl)-1-propylxanthine
- R
- inactive receptor conformation
- R*
- active receptor conformation
- RT-PCR
- reverse-transcription polymerase chain reaction
- SCH 442416
- 2-(2-furanyl)-7-[3-(4-methoxyphenyl)propyl]-7H-pyrazolo[4,3-e][1,2,4]triazolo[1,5-c]pyrimidin-5-amine
- ZM241385
- 4-(2-[7-amino-2-(2-furyl)[1,2,4]triazolo[2,3-a][1,3,5]triazin-5-ylamino]ethyl)phenol
- Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}