


Abstract
The goal of the present study was to investigate a possible role for regulators of G protein-signaling (RGS) proteins in opioid receptor (OR) desensitization using cultured Xenopus laevis dermal melanophores. Morphine-induced pigment aggregation in a melanophore cell line stably expressing the murine μ OR (μOR) was quantified over time. Responses of the μOR (a Gi-linked receptor) exhibited a time-dependent desensitization, which varied with the concentration of morphine used. In contrast, much less desensitization was observed in response to melatonin, effects mediated through the cells’ endogenous melatonin receptor (which is also Gi-linked). To further study OR desensitization, melanophores lacking a μOR were transiently transfected with plasmids encoding the μOR alone or in combination with plasmids encoding one of several RGS subtypes (RGS1, RGS2, RGS3, or RGS4). Overexpression of RGS2, but not the other RGS subtypes, produced a rightward shift in the morphine concentration-response curve. RGS protein overexpression also decreased the magnitude of morphine-induced responses. Finally, the effect of a mutant form of Gαi1, which is insensitive to RGS action, was investigated with respect to its ability to alter the response of the μOR to morphine. Expression of the mutant Gαi1prolonged morphine-induced pigment aggregation and produced leftward shifts in concentration-response curves, compared with expression of wild-type Gαi1. These results demonstrate that specific RGS proteins can dampen signals initiated by agonist activation of the μOR, and support a possible role for RGS proteins in OR desensitization.
The clinical utility of opiates is hampered by the development of tolerance, which can be defined as a progressive decrease in drug potency with repeated administration. An improved understanding of the cellular and molecular mechanisms underlying opiate tolerance would have tremendous clinical implications.
Responses to opiates are mediated through three major classes of opioid receptors (ORs): μ, δ, and κ, all of which couple to the Gαi family of G proteins. The μOR in particular is important for the analgesic and rewarding properties of opiate drugs (Matthes et al., 1996). The availability of cDNAs encoding ORs has opened many avenues of research, including studies of receptor localization, ligand-receptor interactions, receptor structure and function analyses, and receptor desensitization (Mansour et al., 1995). Although much has been learned regarding ORs and their signaling pathways, the mechanisms underlying receptor desensitization and development of drug tolerance remain relatively poorly understood (Nestler and Aghajanian, 1997).
Recently, a family of mammalian proteins with direct effects on the GTPase activities of Gα subunits has been discovered. To date, 19 mammalian regulators of G protein-signaling (RGS) proteins have been described (Dohlman and Thorner, 1997; Gold et al., 1997; Koelle, 1997;Berman and Gilman, 1998). These proteins mediate their physiological effects by facilitating the ability of Gα subunits to hydrolyze bound GTP and thereby increase the rate at which the Gα subunits reassociate with their corresponding Gβγ complexes. In this manner, RGS proteins hasten the return of G proteins to their inactive, GDP-bound state. However, more recent data indicate that RGS proteins can hasten the kinetics of receptor responses (Doupnik et al., 1997;Saitoh et al., 1997). As a result, the net effect of RGS proteins on the functioning of G protein-coupled receptors remains unclear.
The goal of the present study was to investigate a possible role for RGS proteins in μOR desensitization. We employed a bioassay developed to quantitate the functional effects of ligands on G protein-coupled receptors (Lerner, 1994). The bioassay relies on the ability of cultured dermal melanophores from Xenopus laevis to retain in vitro their ability to translocate pigment in response to changes in second messenger levels, including Gαi-mediated responses, induced by activation of specific G protein-coupled receptors. Major advantages of this assay are that it: 1) provides measures of receptor-initiated functional responses over a wide time course (seconds to hours); 2) allows for multiple measurements of the same sample over time because cell sacrifice is unnecessary; 3) does not require ligand modification (e.g., radiolabeling) for response quantitation; and 4) enables the study of a wide range of receptors and intracellular pathways. For these reasons, the assay provides a rapid and high-throughput system to study functional responses of G protein-coupled receptors (Lerner, 1994) and is currently widely used in drug discovery efforts (Jayawickreme and Kost, 1997).
Using the melanophore-based bioassay system, the effects of morphine on the μOR were characterized. The responses of the D2B dopamine receptor (D2R) and the cells’ endogenous melatonin receptor, both of which are also Gαi-coupled (Potenza et al., 1994), were studied for comparison. We investigated the effects of individual RGS proteins, as well as a mutant Gαi subunit defective in its interaction with RGS proteins, on receptor responses. The results demonstrate differential abilities of specific RGS proteins to influence the functional responsiveness of the μOR and other receptors to agonist exposure.
Materials and Methods
Cell Culture.
Xenopus fibroblasts and melanophores were isolated and propagated as described previously (Potenza et al., 1994). Fibroblasts were grown at 27°C in room air in 70% L-15 Medium (Sigma, St. Louis, MO) supplemented with 20% fetal calf serum (Life Technologies, Inc., St. Louis, MO). The melanophores were grown at 27°C in room air in media conditioned for 3 to 4 days by the fibroblasts, collected by passage through a 0.22-μm sterile filter (Falcon, Franklin Lakes, NJ). Media and PBS were diluted with water to 70% strength for all work with the Xenopuslines. Pigment translocation assays were performed in serum-free 70% L-15 to minimize potential exposure of the cells to bioactive moieties from fetal calf serum during the assays. All melanophores used for experimentation stably expressed the human β2-adrenergic receptor (β2AR; Potenza et al., 1992), because the β2AR cell line propagates and electroporates more efficiently than wild-type melanophores (A. Roby-Shemkovitz and M. R. Lerner, personal communication). The stable cell line expressing the μOR was obtained as previously described (Huang, 1996) by cotransfection of the β2AR melanophore cell line with pJGμOR and pcDNAI/NEO (Invitrogen, Carlsbad, CA) and isolation of individual clonal colonies by the process of limiting dilution in the presence of G418.
Microtiter Plate Assays.
Melanophores were seeded into standard 96-well tissue culture plates (Falcon) and grown to approximate confluence (about 7,000–10,000 cells/well). One day before assaying, the medium was removed and the cells were washed with either 70% PBS or L-15. Cells were then incubated overnight in 50 μl of 70% L-15/well. Fifty microliters of fresh 70% L-15 were added to each well and the cells were incubated in room light for 0.5 h (nontransfected μOR/β2AR and β2AR cells) or in room light for 0.5 h and under a 40-W bulb (General Electric, Albany, NY) with light source located 20 inches from the cells to induce pigment dispersion (transiently transfected cells). Fifty microliters of 70% L-15 containing three times the final concentration of test ligand were added to each well at “time zero”, and the cells were maintained in room light (nontransfected μOR/β2AR and β2AR cells, transfected cells for naloxone experiments) or under 40-W bulb light source (transfected cells for (−)quinpirole and RGS experiments) for the duration of the experiment, except during the 10- to 12-s periods during which measurements were being performed by the microtiter plate reader (SLT/Tecan, Hillsborough, NC). The additional photostimulation was generally employed with melanophores transiently transfected with G protein-coupled receptors because D2R-expressing cells are not fully pigment-dispersed under room light alone (Potenza et al., 1994). Morphine and melatonin were obtained from Sigma and (−)quinpirole was obtained from Research Biochemicals International (Natick, MA). All drug preparations were made either on the day of use or obtained from aliquots stored at −20°C. Transmission at 620 nm was measured by a 340 ATTC microtiter plate reader (SLT/Tecan, Hillsborough, NC) using the agglutination mode of the SOFT 2000 program (SLT/Tecan). The agglutination mode acquires 20 separate transmission readings of each well for each time point. In each case, the final transmission reading was discarded (Potenza and Lerner, 1992), and the remaining 19 values were averaged and treated as an individual value. Data stored in ASCII files on a Zeos 386 computer were transferred to a MacIntosh using the Apple File Exchange program for reduction, using Microsoft Excel software. Using the curve-fit application of Kaleidagraph (v3.0), EC50 values were calculated. When curve fitting data, the general equation employed was as follows:
In cases where cells were incubated overnight and then rechallenged with drug, incubation proceeded overnight in the continued presence of white light. Twenty-four hours after initial drug addition, absorbance readings were performed (second time zero) and a fresh aliquot of drug was added. The additional drug was added at a 4-fold concentration in 50 μl of 70% L-15 to the 150 μl of 70% L-15 in the well to yield a working concentration of drug. The initial reading (original time zero) was used as the intrawell standard for calculations of AF/AI − 1 for readings on both days 1 and 2 of the experiments. Microtiter plate experiments were performed as above, and data shown are representative of at least duplicate evaluations.
Statistical analyses of EC50 values were performed using Student’s t tests (n = 3/group). Confidence intervals adjusted for Bonferroni corrections were calculated to determine statistical significance.
Plasmid DNA Constructs.
The plasmid encoding the D2R, pJGD2R, contains the human D2B dopamine receptor under the transcriptional regulation of a cytomeglavirus (CMV) promoter and was constructed as described previously (Potenza et al., 1994). The murine μOR was subcloned into the eukaryotic expression vector pJG3.6 (Potenza et al., 1994) under the transcriptional regulation of the CMV promoter as described (Huang, 1996) to create the plasmid pJGμOR. LacZ-encoding plasmids used in electroporations were either pON260 or pJGLacZ, as described previously (Potenza et al., 1994). RGS-encoding plasmids, containing human cDNA copies of RGS1 (prcCMV-RGS1), RGS2 (prcCMV-RGS2), RGS3 (prcCMV-RGS3), or RGS4 (pCR3-FLAG-RGS4), were generous gifts of John H. Kehrl (National Institutes of Health, Bethesda, MD) and used for electroporation without modification (Neill et al., 1997). Wild-type (H6pQE6Gia1, H6pQE6Go) and mutant (H6pQE6Gia1G184S, H6pQE6GoG184S) plasmid constructs of Gαi1 and Gαo, respectively, were generous gifts of Keng-Li Lan and Richard R. Neubig (University of Michigan, Ann Arbor, MI; Lan et al., 1998). Wild-type (αqEE2pcDNAIAmpa) and mutant (αqEE2GSpcDNAIAmpa) plasmid constructs of Gαq were generous gifts of Donald Apanovitch and Henrik G. Dohlman (Yale University, New Haven, CT; DiBello et al., 1998). The coding sequences for the wild-type and mutant Gαi1 and Gαo subunits were excised from their respective plasmids and placed under the transcriptional regulation of a CMV promoter in JG3.6 using standard protocols. During the plasmid constructions, the sequences coding for the histidine tags were removed, generating wild-type and mutant Gαi1 and Gαo plasmids lacking sequences encoding tags. The identities of all constructs were verified by restriction enzyme digest analysis with enzymes purchased from Boehringer-Mannheim (Indianapolis, IN) or New England Biolabs (Beverly, MA). Plasmid DNA amplification was accomplished using mini-prep and endo-free maxi-prep protocols (Qiagen, Santa Clarita, CA).
Transfections.
Plasmid DNA was transfected into the melanophores by the method of electroporation using an Electo Cell Manipulator 600 (BTX, San Diego, CA) at settings of 575 μF, 475 V, and R10 as described elsewhere (Potenza et al., 1994). Briefly, cells were washed with 70% PBS and detached by trypsinization (2.5% trypsin/3 mM EDTA; Life Technologies, Inc.) in 70% PBS. After inactivating the trypsin with serum (Life Technologies, Inc.) and collecting the cells by centrifugation at 2,000g at 4°C for 5 min, the supernatant was removed and the cell pellet resuspended in 5 ml of ice-cold 70% PBS and recentrifuged as above. After removing the supernatant, the cells were resuspended in ice-cold 70% PBS at a density of 5,000,000 cells/ml. Twenty micrograms of plasmid DNA were added to 400 μl of cells and the resulting mix was incubated on ice for 10 min in a prechilled electroporation cuvette (0.2 cm gap; BTX, San Diego, CA). The cuvette contents were triturated with a micropipetor just before electroporation and plated in media immediately after electroporation. Cells were plated onto tissue culture ware pretreated with Cell-Tak (Colloborative Research, Bedford, MA) to maximize survival of transfected cells following electroporation. Cells were assayed 3 days after electroporation, as described above.
Transfection efficiency was routinely assayed by in situ staining for β-galactosidase activity of cells transfected with a lacZ-encoding plasmid, according to published procedures (Potenza et al., 1994). Transfection efficiency, as determined by this method, did not differ significantly among the samples on given days. Using the above-described procedure, transfection efficiencies of 30 to 60% were reliably observed. D2R- and μOR-expressing melanophores were generated by transfecting with a 1:4 weight-to-weight mixture of the pJGD2R and pJGμOR, respectively, and lacZ-encoding pJGlacZ or pON260. In the time-course analysis of (−)quinpirole on D2R-expressing melanophores (Fig. 1E, 1F), a 1:50 weight-to-weight mixture of pJGD2R and pJGlacZ was used. During cotransfections with plasmids encoding receptors and RGS proteins, a 1:1:3 weight-to-weight mixture of receptor/RGS/LacZ-encoding plasmids was employed. In some experiments, comparable levels of expression of the various RGS proteins were confirmed by Northern blotting by use of published methods (Gold et al., 1997). LacZ-transfected (“mock-transfected”) cells were produced by electroporation of the melanophores with 20 μg of either pJGlacZ or pON260. For experiments investigating Gα subunit function, 8 μg of Gα-encoding and 12 μg of LacZ-encoding plasmids were used per electroporation into μOR-expressing melanophores.

Time-course and concentration-response analyses of μOR- or D2R-expressing melanophores in response to morphine, melatonin, or (−)quinpirole. μOR-expressing and control cells were incubated in the continuing presence of morphine (A and B) or melatonin (C and D), or D2R-expressing and control cells were incubated in the continuing presence of (−)quinpirole (E and F). Changes in absorbance through the cells were measured and used to quantitate receptor-mediated pigment translocation as described in the text. Δ(AF/AI − 1), a measure of pigment aggregation, is defined as the response ratio (seeMaterials and Methods). Error bars represent S.E. values of the mean for triplicate samples. For graphs of time-course analyses, concentrations of indicated drugs are displayed in the figure insets. For graphs of concentration-response curves, cell types used are displayed in the figure insets. Results shown are representative of at least duplicate experiments.
Results
Morphine-Induced Pigment Aggregation in a Melanophore Cell Line Stably Expressing Murine μOR.
As a first step in examining μOR function, the properties of a stable cell line of melanophores expressing the murine μOR were explored. After pretreating cells with light to induce pigment dispersion, cells were exposed to varying concentrations of morphine in the continued presence of light, and pigment aggregation was quantitated over time by determination of absorbance changes through the cells (Fig. 1). Increasing amounts of pigment aggregation, as is observed after stimulation of Gαi-coupled receptors in the melanophores (Potenza et al., 1994), results in decreasing magnitudes of absorbance readings and negative values of the response ratio Δ(AF/AI − 1) of increasing magnitude. As expected, increasing concentrations of morphine produced successively increasing amounts of pigment aggregation in the cells (Fig. 1A). The response to morphine appeared within physiologically relevant concentrations, with maximal functional responses seen at 1 and 10 μM. Over the 7-h time course of the experiment, the responses to morphine diminished. The intermediate concentration of 100 nM produced a response peaking at 30 min and returning to baseline over the 7-h time course of the experiment. The higher concentrations of 1 and 10 μM produced responses peaking by 30 min, remaining maximal for an additional 90 min, and then, in a superimposable fashion, returning toward baseline thereafter. The EC50 value for morphine on the cells was 91.4 ± 4.3 nM at 30 min (Fig. 1B).
The response to morphine mediated by the murine μOR can be compared with that of melatonin mediated by the melatonin receptor endogenous to these melanophores (Potenza et al., 1994). The μOR-expressing melanophores aggregated their pigment in a concentration-dependent fashion in response to melatonin (Fig. 1C). Time-course analysis of the melatonin response shows a pattern that differs from that seen with morphine. For example, whereas the cells exposed to maximal concentrations of morphine (1 and 10 μM) showed a significant decrease from the maximal response by 7 h, those exposed to maximal concentrations of melatonin (10 and 100 nM) did not. The EC50 value at 30 min for melatonin on the μOR-expressing cells was 56.2 ± 10.1 and 62.8 ± 4.2 nM on the control cells (Fig. 1D).
For comparison, we next investigated the time course of agonist-induced pigment aggregation mediated by another Gi-linked receptor, the human D2R. In a manner similar to μOR responses to high concentrations of morphine, high concentrations of the D2R agonist (−)quinpirole induced pigment aggregation, which returned toward baseline values over the 7-h course of the experiment (Fig. 1, E and F). The EC50 value at 30 min for quinpirole on the D2R-transfected cells was 17.0 ± 3.5 nM. In contrast to the time-course experiments investigating the μOR and melatonin receptor responses, those exploring the D2R response were performed with cells transiently expressing the receptor, because attempts to generate melanophores stably expressing the D2R have not been successful to date (M. N. Potenza and M. R. Lerner, unpublished results). Given the transient nature of D2R expression, we cannot rule out the possibility that response dimunition over the 7-h time course of the experiment is in part mediated by receptor loss. However, this would not appear to be the case, because (−)quinpirole can produce robust responses on initial exposure in cultures at these longer times after transfection (see Potenza et al., 1994).
Antagonism of Morphine-Induced Pigment Aggregation by Naloxone.
To study further the characteristics of μOR responses in the cultured melanophores, the potential for naloxone, a specific μOR antagonist, to inhibit morphine-induced pigment aggregation was investigated (Fig. 2, A and B). Cells transiently transfected with μOR-encoding plasmid were pretreated with light to induce pigment dispersion and then exposed to 100 nM morphine and varying concentrations of naloxone. Naloxone blocked the morphine-induced pigment aggregation in a dose-dependent manner. These findings confirm that morphine’s effects on the melanophores are mediated via the μOR.

Time-course and concentration-response analyses of naloxone-mediated antagonism of morphine-induced pigment aggregation in μOR-expressing melanophores. Pigment aggregation over time (A) and at 30 min (B) in response to 100 nM morphine and varying concentrations of the μOR antagonist naloxone was measured in cells transfected with μOR-encoding plasmid. LacZ-transfected cells did not show appreciable pigment aggregation in response to morphine. Changes in absorbance through the cells were measured and used to quantitate receptor-mediated pigment translocation as described in the text. Δ(AF/AI − 1), a measure of pigment aggregation, is defined as the response ratio (see Materials and Methods). Error bars represent S.E. values of the mean for triplicate samples. For the time-course graph, concentrations of naloxone are displayed in the figure inset. For the concentration-response graph, cell types used are displayed in the figure inset. Results shown are representative of at least duplicate experiments.
Rechallenge of μOR-Expressing Cells to Morphine and Melatonin.
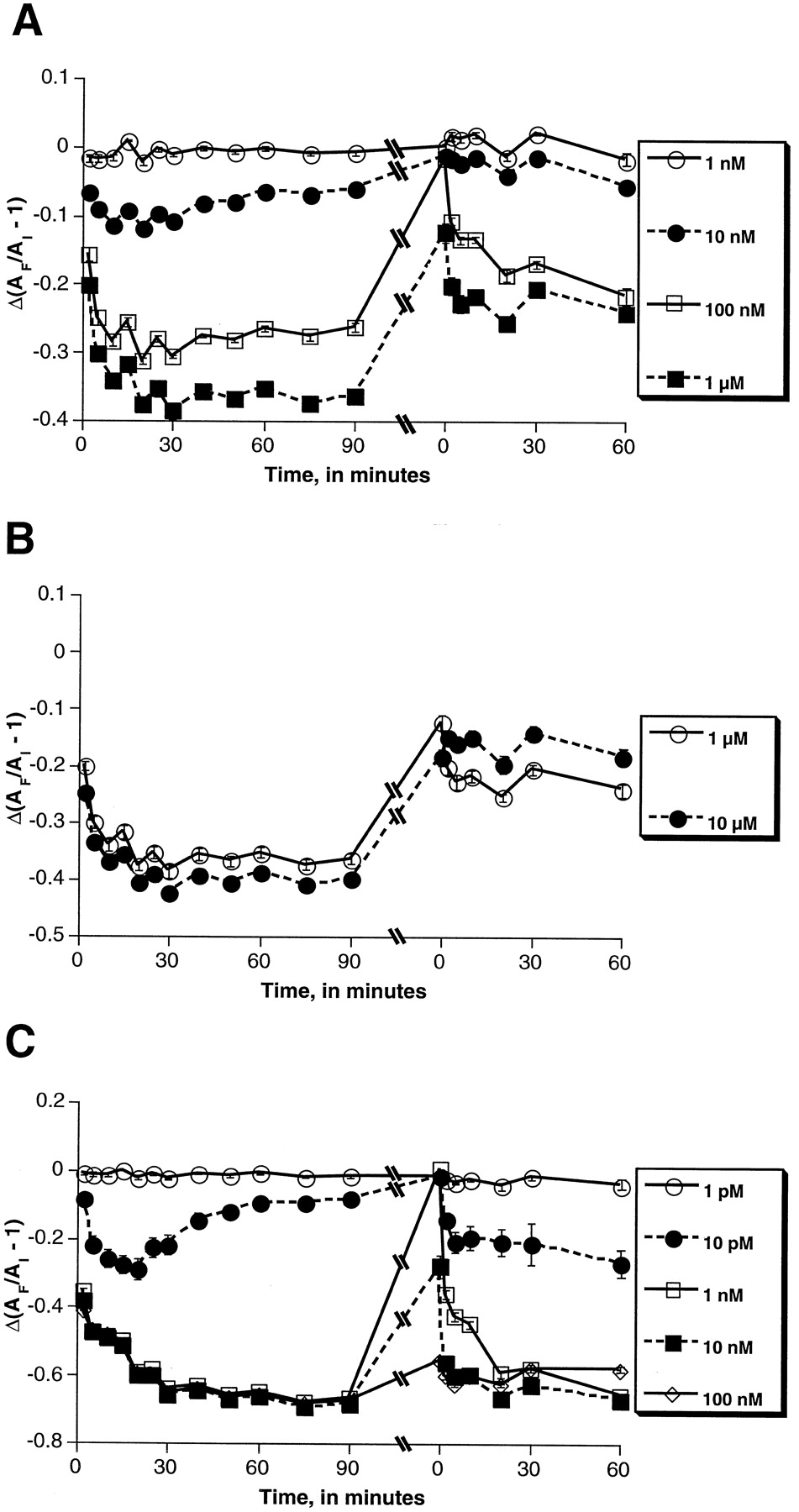
To investigate further possible desensitization in μOR responses, cells were exposed to morphine or melatonin and then rechallenged with the same drug (Fig. 3). Incubation proceeded overnight and rechallenge with the same concentration of fresh drug was made (second time zero in Fig. 3) as described in Materials and Methods. Over the 60-min period after the second application of morphine, the drug produced a less robust response for all of the concentrations tested (Fig. 3, A and B). It appears that the higher the concentration of morphine initially applied, the less robust a response was seen on restimulation. For example, re-exposure to 10 μM morphine produced virtually no response (Fig. 3B), whereas re-exposure to 100 nM morphine produced a response approximately two-thirds of that seen initially (Fig. 2A).

Rechallenge of morphine- and melatonin-treated μOR-expressing melanophores with additional drug. Cells were incubated overnight in the continued presence of morphine (A and B) or melatonin (C) and rechallenged 24 h later with the same concentration of the same drug to which they were originally exposed. Changes in absorbance through the cells were measured and used to quantitate receptor-mediated pigment translocation as described in the text. Δ(AF/AI − 1), a measure of pigment aggregation, is defined as the response ratio. The initial reading (original time zero) was used as the intrawell standard for calculations of AF/AI − 1 for readings on both days 1 and 2 of the experiments, as described in greater detail inMaterials and Methods. Error bars represent S.E. values of the mean for triplicate samples. Concentrations of indicated drugs are displayed in the figure inset for each graph. Results shown are representative of at least duplicate experiments.
Similar experiments were performed to investigate the response of the μOR-expressing melanophores to melatonin (Fig. 3C). In a fashion similar to the morphine-treated cells, cells exposed to high concentrations of melatonin (100 nM and 10 nM) did not return to the pretreatment state of pigment dispersion. However, in contrast to the morphine-treated cells, rechallenge with melatonin at all concentrations tested produced responses that closely approximated those seen initially. Low and intermediate concentrations of melatonin (10 pM and 1 nM) resulted in pigment aggregation that returned to baseline over 24 h. The inability of these low concentrations to maintain pigment aggregation might be accounted for by a pattern of receptor desensitization differing from that of the μOR. However, it is also possible that other factors may be involved: for example, at lower concentrations, the melatonin may have degraded in the medium, or that the melatonin-induced, Gi-linked stimulus is not of sufficient magnitude to offset continued cAMP accumulation from photostimulation.
Effect of RGS Proteins on Responses to Morphine in Transiently Transfected Melanophores Expressing μOR.
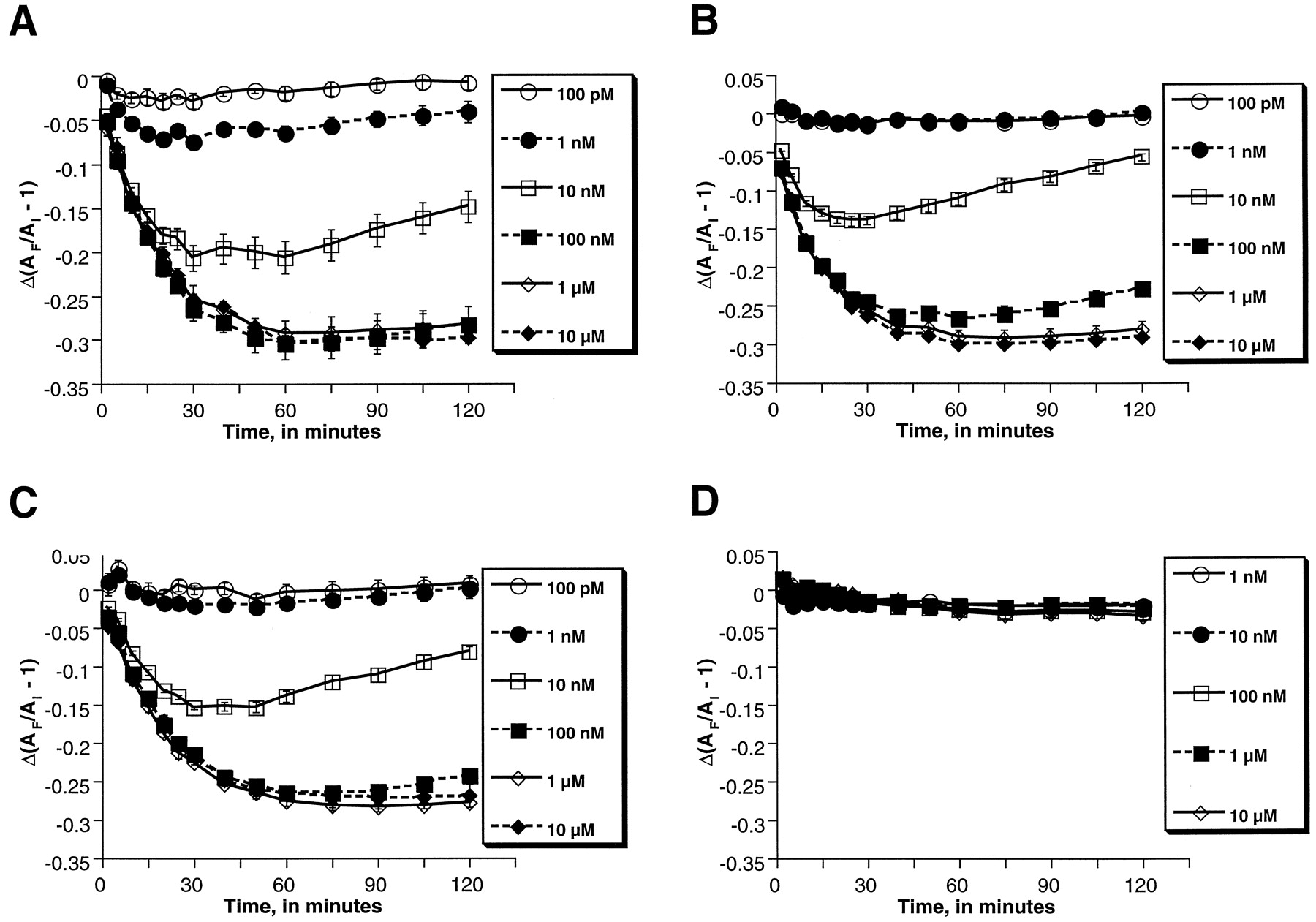
To investigate the effect of specific RGS proteins on functional responses mediated by the μOR, melanophores were transiently transfected with a plasmid encoding the μOR alone or in combination with plasmids encoding RGS1, RGS2, RGS3, or RGS4. Transient transfection techniques were employed with the goal of targeting the μOR- and RGS-encoding plasmids to the same subset of cells. Cells were also cotransfected with a plasmid encoding LacZ to control for transfection efficiency by staining for β-galactosidase activity. Cells were tested over time for their responses to various concentrations of morphine (Fig.4). Time-course analysis over the 2-h time course of the experiment revealed that cells transfected with the RGS-encoding plasmids retained their abilities to translocate pigment in response to morphine. However, certain RGS proteins, particularly RGS2, were able to diminish the sensitivity of the cells to morphine, as is most evident from comparison of the cells’ responses to intermediate concentrations (10 and 100 nM) of morphine (Fig. 4, A versus B). Determination of EC50 values at multiple time points revealed the greatest changes with expression of RGS2, which resulted in approximately 2-fold shifts to the right in EC50 values (Table 1). No consistent effect of the other RGS subtypes was observed. The differential effects of RGS2 and the other RGS proteins on μOR responses did not seem to be caused by differences in the levels of expression of the transfected RGS proteins: Northern blotting revealed comparable levels of expression of each of the RGS mRNAs (data not shown).

Time-course analyses of morphine-induced pigment aggregation in melanophores cotransfected with plasmids encoding the μOR and RGS2. Pigment aggregation in response to morphine was measured in cells transfected with plasmids encoding the μOR alone (Fig. 4A) or in combination with one encoding RGS2 (B). LacZ-transfected cells did not show appreciable pigment aggregation in response to morphine (C). Δ(AF/AI − 1), a measure of pigment aggregation, is defined as the response ratio (seeMaterials and Methods). Error bars represent S.E. values of the mean for triplicate samples. Concentrations of morphine used are displayed in the figure inset for each graph. Results shown are representative of at least duplicate experiments.
EC50 values for morphine on μOR/RGS-expressing cells
Differences were also noted in the absolute magnitude of the response ratio in RGS-transfected melanophores. This was seen not only with RGS2 (as shown in Fig. 4), but also for the other RGS proteins as well (not shown). The response ratio can be influenced by multiple factors, including plating density and, in transiently transfected cells, electroporation efficiency. Electroporation efficiency, as assessed by β-galactosidase staining, did not appear to account for these differences. To investigate the potential contribution of plating density, μOR/RGS-transfected melanophores from the identical electroporations tested with morphine were plated at the same density and exposed to varying concentrations of melatonin. Samples with lesser degrees of morphine-induced pigment aggregation did not correlate with those displaying lesser degrees of melatonin-induced pigment aggregation, as assessed by response ratio values (not shown). Moreover, the EC50 values of melatonin on μOR/RGS-expressing cells were not significantly different from μOR-expressing or mock-transfected cells (Table2). The lack of change in the EC50 values for melatonin observed with RGS protein expression should be interpreted cautiously, given that expression of the melatonin receptor was not targeted to the same subset of cells expressing the RGS proteins, as was the case with the μOR. That is, given that the melatonin receptor is endogenously expressed in the melanophores, it is expected that the transient transfection techniques used would result in a lower percentage of the melatonin receptor-expressing cells to express an RGS protein, compared with the μOR-expressing cells, making it potentially more difficult to observe an effect on the response to melatonin. Finally, it is conceivable that the different effects of the RGS proteins on the μOR and melatonin receptor might involve differences in the patterns of desensitization of the two receptors, although this remains speculative.
EC50 values for melatonin on μOR/RGS-expressing cells
Effect of RGS Proteins on Response to (−)Quinpirole in Transiently Transfected Melanophores Expressing D2R.
To investigate the effect of RGS proteins on another Gαi-coupled receptor, the D2R was expressed transiently alone and in combination with various RGS proteins and tested with the D2R agonist (−)quinpirole (Fig.5). D2R-expressing cells, in the absence or presence of RGS proteins, showed the ability to aggregate pigment in response to (–)quinpirole, whereas LacZ-transfected cells did not. As seen with morphine on the μOR, RGS proteins influenced the D2R-mediated functional response to (−)quinpirole, although different subtypes of RGS proteins were most effective for the two receptors. With the D2R, RGS3 produced the greatest decrement in (−)quinpirole response, as is most evident from comparison of the cells’ responses to intermediate concentrations (1, 10, and 100 nM) of the drug (Fig. 5, A versus B).

Time-course analyses of (−)quinpirole-induced pigment aggregation in melanophores cotransfected with plasmids encoding the D2R and RGS3 or RGS2. Pigment aggregation in response to (−)quinpirole was measured in cells transfected with plasmids encoding the D2R alone (A) or in combination with those encoding RGS3 (B) or RGS2 (C). LacZ-transfected cells did not show appreciable pigment aggregation in response to (−)quinpirole (D). Δ(AF/AI − 1), a measure of pigment aggregation, is defined as the response ratio (see Materials and Methods). Error bars represent S.E. values of the mean for triplicate samples. Concentrations of (−)quinpirole used are displayed in the figure inset for each graph. Results shown are representative of at least duplicate experiments.
RGS3 also had the greatest effect on EC50 values for quinpirole on the D2R (Table 3). D2R/RGS3-transfected cells displayed EC50 values more than 3-fold greater than the corresponding ones for D2R-transfected cells at the 30-, 60-, and 120-min time points. The other RGS proteins tended to produce similar effects, but these were smaller in magnitude, compared with those for RGS3. For example, RGS2 diminished the response to (−)quinpirole (Fig. 5, A versus C) and produced an approximately 2-fold shift in the EC50 value for (−)quinpirole (Table 3).
EC50 values for (−)quinpirole on D2R/RGS-expressing cells
Effect of Wild-Type and Mutant Gαi1 Subunits on μOR Function.
To investigate further the effect of RGS proteins on μOR-mediated responses, wild-type or mutant forms of Gαi1 were transfected into the μOR-expressing melanophores. The mutant version contains a glycine to serine substitution at position 183 that markedly lowers its affinity for RGS proteins and makes it virtually unresponsive to stimulation by RGS proteins (DiBello et al., 1998; Lan et al., 1998). As shown in Fig.6, overexpression of the mutant Gαi1 potentiated responses of the μOR to morphine. Such increased and prolonged responses to morphine were most evident at intermediate concentrations of drug (e.g., 10 and 31.6 nM) in the cells expressing the mutant protein, compared with those expressing the wild-type version (Fig. 6, A versus B). EC50 values of morphine on the melanophores expressing the mutated version of Gαi1 were consistently one-half to two-thirds that observed in cells expressing the wild-type form over the time course of the experiment (Table 4). In contrast to the results obtained with the Gαi1 constructs, similar experiments performed with the wild-type and corresponding glycine to serine mutant versions of Gαo and Gαq found minimal differences in morphine responses between cells transfected with the wild-type and mutant forms of the proteins (not shown).

Time-course analyses of morphine-induced pigment aggregation in melanophores stably expressing the μOR and transiently transfected with plasmids encoding wild-type or mutant Gαi1 proteins. Pigment aggregation in response to morphine was measured in μOR-expressing cells transiently transfected with plasmids encoding wild-type (A) or mutant (B) versions of Gαi1. Δ(AF/AI − 1), a measure of pigment aggregation, is defined as the response ratio (seeMaterials and Methods). Error bars represent S.E.s of the mean for triplicate samples. Concentrations of morphine are listed in the figure insets. Results shown are representative of at least duplicate experiments.
EC50 values for morphine on μOR/Gαi1-expressing cells
Discussion
In this series of experiments, we investigated the regulation of agonist-induced responses over time for the μOR. Study of the μOR was facilitated by the creation of a stable melanophore cell line expressing this receptor. The inability of even high concentrations of morphine to sustain a maximal response over the course of hours in the μOR stable line suggested the possibility of homologous desensitization, consistent with findings in other cell systems (Zimprich et al., 1995). The finding of decreasing responsivity observed on rechallenge with morphine provides additional evidence consistent with such desensitization. Given that similar studies investigating the effects of melatonin on the endogenous Gαi-coupled melatonin receptor showed a different pattern of response over hours and on drug rechallenge suggests receptor specificity in the responses. Moreover, given that the μOR is not under the transcriptional regulation of its endogenous promoter, it is likely that information necessary for the observed pattern of desensitization lies within the receptor protein itself.
We next sought to investigate the effects of specific RGS proteins in regulating functional responses of the μOR. RGS proteins have been shown to increase the rates of hydrolysis of GTP to GDP for many types of Gα subunits, including Gαi subtypes, thereby facilitating their return to an inactive state. We investigated the abilities of four individual RGS proteins, RGS1–4, to influence responses of the μOR to morphine. Three of the four RGS proteins studied (RGS2–4) have been reported to be expressed in brain regions (e.g., striatum) of high μOR and D2R expression (Burchett et al., 1998). One of these proteins, RGS2, was found to have a greater effect on dampening morphine-induced μOR activation than the other RGS subtypes. In contrast, a different RGS protein, RGS3, was found to have relatively greater effects on D2R responses to (−)quinpirole, with RGS2 displaying somewhat less of an effect. Responses of the melatonin receptor were not consistently affected by any of the RGS proteins examined. These findings raise the interesting possibility that some degree of RGS/receptor specificity might exist in the regulation of receptor responses.
The basis for this differential action of the RGS proteins on the μOR, D2R, and melatonin receptor remains unknown. All three receptors couple to the Gαi family of G proteins, which raises the possibility that perhaps the receptors act through distinct subtypes, or at least distinct cellular pools, of these proteins. Support for selective actions of various RGS proteins on a given receptor comes from several recent studies. In one report, RGS3 displayed a greater ability, compared with RGS1, RGS2, and RGS4, to terminate physiological responses to gonadotropin-releasing hormone in an in vitro assay (Neill et al., 1997) and, in a separate study, RGS1, RGS3, and RGS4, but not RGS2, dramatically accelerated agonist activation of muscarinic m2 or serotonin 1A receptors (Doupnik et al., 1997). More recently, the N-terminal domain of RGS4 was demonstrated to confer both high affinity and receptor selectivity to the protein (Zeng et al., 1998). These findings support the hypothesis that individual RGS proteins may preferentially interact directly with specific receptor-G protein complexes. A related question is whether specific RGS proteins exert differential effects on specific subtypes of G protein α subunits. However, despite the identification of cDNAs corresponding to 19 mammalian RGS proteins, it has been difficult to demonstrate high levels of specificity of any given RGS protein for a particular subtype of α subunit, with most proteins examined able to regulate both Gαi and Gαq subunits (see Berman and Gilman, 1998).
We also show in the present study that mutant forms of Gαi1 can potentiate functional responses of the μOR. The mutant Gαi1 is unresponsive to RGS proteins; that is, RGS proteins are unable to activate the GTPase activity of the α subunit (Lan et al., 1998). As a result, the mutant Gαi1 functions in a sense as a dominant negative mutation for RGS proteins. Our findings, therefore, are consistent with the possibility that the desensitization observed in μOR responses in the cultured melanophores under normal conditions (e.g., Fig. 1) is dependent, in part, on RGS function. Thus, we show in this study not only that RGS proteins can promote diminution of the μOR response, but also that RGS proteins are required for the diminution normally seen in the μOR response in the melanophore system. Although the specific site of action of the RGS proteins on pigment translocation within the melanophores is not known with absolute certainty and could conceivably involve effects on downstream proteins, all available data support the notion that RGS proteins directly affect the function of specific G proteins and the immediate proteins (i.e., G protein-coupled receptors) with which they interact (Berman and Gilman, 1998; Zeng et al., 1998).
RGS proteins represent only one of several potential levels at which the functional responses of G protein-coupled receptors can be regulated. G protein-coupled receptor kinases (GRKs) can phosphorylate specific residues in the receptors that causes their association with arrestins and prevents subsequent receptor activation of G protein α subunits. Specific GRKs have been shown to phosphorylate and desensitize ORs in vitro (Pei et al., 1995; Kovoor et al., 1997; Zhang et al., 1998), and chronic morphine administration has been shown to up-regulate one subtype of GRK in specific brain regions in vivo (Terwilliger et al., 1994), consistent with a possible role for GRK-mediated phosphorylation in OR tolerance. ORs have also been shown to be phosphorylated by other protein kinases, which could result in altered receptor sensitivity as well (e.g., see Koch et al., 1997;Chakrabarti et al., 1998). Finally, it is known that agonists can induce internalization of ORs (which may be a phosphorylation-dependent process; Keith et al., 1998), as well as changes in levels of receptor expression (see Blake et al., 1997; Afify et al., 1998), both in vitro and in vivo. Results of the present study introduce another level—RGS proteins—at which agonist-initiated responses of ORs (and other G protein-coupled receptors) can be regulated. A major question for future research is to determine the relative contribution of each of these mechanisms to the OR desensitization and tolerance that occurs in specific neuronal cell types in vivo under various physiological and pharmacological conditions.
The ability of specific RGS proteins to modify responses to the μOR and D2R can begin to be understood within a functional context. Both receptor types are expressed at high levels in striatal regions of brain (Baik et al., 1995; Mansour et al., 1995), where they have been implicated in mediating the rewarding properties of opiates, psychostimulants (e.g., cocaine and amphetamine), and other drugs of abuse (Harris and Aston-Jones, 1994; Matthes et al., 1996; Nestler and Aghajanian, 1997). Recently, Burchett et al. (1998) reported the relative levels of specific RGS mRNAs in striatum. Interestingly, RGS2 mRNA was found to be most abundant of the several RGS subtypes examined, whereas levels of RGS3 mRNA were relatively low. The same study also reported that acute administration of amphetamine increased levels of RGS2 and RGS3 mRNA in this brain region. In another recent study, RGS2 mRNA was found to be highly regulated in striatum by electrical activity as well as by dopamine-acting agents (Ingi et al., 1998). Results of the present study indicate that such changes in levels of expression of RGS2 and RGS3 might be expected to result in alterations in the functional responses of μOR, D2R, and perhaps other G protein-coupled receptors in this brain region.
Additional work is now needed to directly demonstrate effects of RGS proteins on receptor function in vivo and to study the consequences of such regulation with respect to various behavioral phenomena. Nevertheless, results of the present study support a scheme wherein changes in the levels or activity of RGS proteins regulate the functional responsiveness of G protein-coupled receptors. The results also support the possibility (Gold et al., 1997) that specific RGS subtypes, because of their regional and functional selectivity, represent potential targets for the development of novel psychotherapeutic agents.
Acknowledgments
We thank Richard R. Neubig and Henrik G. Dohlman for helpful comments during preparation of the manuscript.
Footnotes
-
Send reprint requests to: Eric J. Nestler, Connecticut Mental Health Center, 34 Park St., New Haven, CT 06508. E-mail:eric.nestler{at}yale.edu
-
↵1 This research was supported by grants from the National Institute on Drug Abuse (to E.J.N.) and the National Institute of Mental Health (to E.J.N.), a Young Investigator Award from the National Alliance for Research in Schizophrenia and Depression (to M.N.P.), a Biological Sciences Training Program training grant (to M.N.P.), and by the Connecticut Mental Health Center. The research presented in this publication was recognized with a Lilly/APA Resident Research Award (to M.N.P.).
- Abbreviations:
- OR
- opioid receptor
- RGS
- regulators of G protein signaling
- β2AR
- β2-adrenergic receptor
- D2R
- D2B dopamine receptor
- GRKs
- G protein-coupled receptor kinases
- μOR
- μ opioid receptor
- Received January 28, 1999.
- Accepted June 15, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}