


Abstract
Although there is strong evidence to suggest that flavonoid consumption is beneficial to human health, the extent to which flavonoids are absorbed and the mechanisms involved are controversial. Contrary to common dogma, we previously demonstrated that quercetin 4′-β-glucoside, the predominant form of the most abundant dietary flavonoid, quercetin, was not absorbed across Caco-2 cell monolayers. The aim of this study was to test the hypothesis that a specific efflux transporter is responsible for this lack of absorption. Transport of quercetin 4′-β-glucoside, alone or with inhibitors, was examined with Caco-2 cell monolayers. In addition, subcellular localization of the multidrug resistance-associated proteins MRP1 and MRP2 was examined by immunofluorescent confocal microscopy. Efflux of quercetin 4′-β-glucoside, a saturable process, was not altered by verapamil, a P-glycoprotein inhibitor, but was competitively inhibited by MK-571, an MRP inhibitor. These data in combination with immunofluorescent localization of MRP2 to the apical membrane support a role for MRP2 in the intestinal transcellular efflux of quercetin 4′-β-glucoside. These results suggest a role for MRP2 in the transport of a new class of agents, dietary glucosides.
Flavonoids are highly diverse, low molecular weight polyphenolic compounds. Due to their ubiquitous distribution in plants, humans are constantly exposed to a large variety of flavonoids, but precise exposure levels are not yet known. Although early estimates reported the average intake of all flavonoids to be as high as 1 g/day (Pierpoint, 1986), newer data suggest that the daily intake may be lower. Based on data from The Netherlands, a Western diet contains approximately 16 mg/day of the most commonly consumed flavonoid, quercetin (Hertog et al., 1993). However, very large interindividual variability can be expected based on dietary preferences. Significant sources of quercetin include onions, apples, tea, and red wines.
Epidemiological data have demonstrated an association between a diet that is rich in quercetin and a significant reduction in the risk of mortality from coronary heart disease (Hertog et al., 1993; Knekt et al., 1996) and a reduced risk of stroke (Keli et al., 1996). Quercetin is a potent antioxidant, which chelates metal ions to prevent the Fenton reaction and is capable of scavenging hydroxyl and peroxy radicals (Manach et al., 1996). In addition, quercetin and quercetin monoglucosides have been shown to inhibit 15-lipoxygenase, an enzyme thought to play a role in the oxidative modification of low density lipoprotein, leading to foam cell formation in the early development of atherosclerosis (da Silva et al., 1998).
Epidemiological studies also support an association between dietary flavonoids and a reduced risk of certain cancers, including stomach carcinoma and lung cancer (Dorant et al., 1996; Knekt et al., 1997). Animal studies (Deschner et al., 1993) as well as in vitro studies suggest that the flavonoids exert preventive effects in cancer. Proposed mechanisms for anticancer benefits include numerous effects on signal transduction pathways involved in cell proliferation (Weber et al., 1996; Lepley and Pelling, 1997) and angiogenesis (Fotsis et al., 1997), as well as inhibition of enzymes involved with procarcinogen bioactivation such as cytochrome P450 (Tsyrlov et al., 1994) and sulfotransferase enzymes (Walle et al., 1995).
Although there is strong evidence to suggest beneficial effects of flavonoids in human health, the extent and mechanism by which flavonoids reach the systemic circulation from dietary sources are controversial. Plant flavonoids are predominantly found as β-glycosides with flavonols (including quercetin) existing as 3, 7, and 4′ O-glycosides, whereas other flavonoids, such as flavones, flavonones, and isoflavones, are mainly glycosylated at position 7 (Price et al., 1997; Fossen et al., 1998). With the notable exception of fermentation and autolysis, which release the aglycone, flavonoid glycosides are relatively resistant to most food preparation methods (Coward et al., 1993). Thus, when consumed, flavonoids are present primarily as glycosides.
An original model of flavonoid bioavailability assumed that flavonoid glycosides were too polar to be absorbed from the small intestine and that absorption was dependent on the cleavage of the β-glycoside linkage by the colonic microflora (Griffiths and Barrow, 1972). Hollman et al. (1995) indirectly calculated the absorption of quercetin aglycone as well as quercetin glucosides from an onion meal in ileostomy patients. Based on the results, they proposed that quercetin glucosides were actively absorbed via the intestinal glucose transporter. In a direct examination of the transcellular absorption of quercetin glucosides with the Caco-2 cell model of human intestinal absorption, in which glucose is very rapidly absorbed, we found that quercetin 4′-β-glucoside was not absorbed, whereas quercetin 3,4′-β-diglucoside demonstrated minimal absorption (Walgren et al., 1998). In fact, contrary to all hypotheses, both of these glucosides demonstrated significant efflux, suggesting that a drug efflux pump may be involved in the basolateral-to-apical transport of these dietary components.
In this study, we have examined the role of known intestinal efflux pumps in the transcellular efflux of quercetin 4′-β-glucoside in human intestinal Caco-2 cell monolayers. Our observations indicate that the multidrug resistance-associated protein MRP2 plays a major role in this efflux. MRP2 identification and subcellular localization to the apical membrane in the Caco-2 cells were established by immunofluorescent confocal microscopy.
Materials and Methods
MK-571 was a generous gift from A. W. Ford-Hutchinson, Merck-Frosst Center for Therapeutic Research, Pointe Claire-Dorval, Quebec, Canada. Dulbecco's PBS with 0.1 g l−1 calcium chloride was purchased from Life Technologies (Grand Island, NY). Quercetin 4′-β-glucoside was isolated from the red onion as described previously (Walgren et al., 1998). Verapamil HCl was obtained from Knoll Pharmaceutical (Whippany, NJ). Anti-human MRP1 and MRP2 antibodies were purchased from Kamiya Biomedical (Seattle, WA). Except where noted, all other chemicals were purchased from Sigma Chemical Co. (St. Louis, MO).
Cell Culture.
Caco-2 cells obtained from American Type Culture Collection (Rockville, MD) were cultured in Eagle's minimum essential medium (MEM) (Cellgro; Mediatech, Herndon, VA) supplemented with 1% MEM nonessential amino acids (Mediatech), 10% fetal bovine serum (Summit Biotechnology, Fort Collins, CO), 100 U ml−1 penicillin, and 0.1 mg ml−1 streptomycin and were grown in a humidified atmosphere of 5% CO2 at 37°C. Cells were subcultured at 80% confluence.
Transport Studies.
For all transcellular transport studies, Caco-2 cells were seeded in 12-mm i.d. Transwell inserts (polycarbonate membrane, 0.4-μm pore size; Corning Costar Corp., Cambridge, MA) in 12-well plates at a density of 1.0 × 105cells cm−2. The basolateral (serosal) and apical (mucosal) compartments contained 1.5 and 0.5 ml of culture medium, respectively. Culture medium was replaced three times a week for 14 days and daily thereafter. Caco-2 cells in Transwells at passage 48 to 93 were used for transport experiments 18 to 25 days post seeding. Inserts with transepithelial electrical resistance (TEER) values >350 Ω cm2 in culture medium were washed twice for 30 min with warm PBS.
Stock solutions of quercetin 4′-β-glucoside in ethanol were diluted with PBS before transport experiments. The resulting maximum final concentration of ethanol, 0.5%, did not affect TEER values or the transport of mannitol, a marker of paracellular transport. All other compounds were dissolved in transport medium. Transport medium containing substrate was added to either the apical (0.5 ml) or basolateral (1.5 ml) side of the inserts, whereas the receiving chamber contained the corresponding volume of PBS. Where applicable, inhibitors (MK-571, verapamil) were added to both chambers. Upon termination of the 1-h incubation at 37°C, samples were collected for immediate analysis.
Samples containing quercetin 4′-β-glucoside were quantified by reversed phase HPLC on a Millennium HPLC system (Waters Corp., Milford, MA) with a Symmetry C18 column, 3.9 × 150 mm, and a model 996 photodiode array detector. The mobile phase consisted of 35% methanol in 5% acetic acid with a flow rate of 0.9 ml min−1. Quercetin 4′-β-glucoside peak areas were measured at 370 nm.
Calculations and Statistics.
Apparent permeability coefficients (Papp) were calculated with the following equation:
Transport data are expressed as a mean flux of five or more determinations ± S.E. ANOVA was used to evaluate differences in flux. A P value <.05 was considered significant. Apparent kinetic constants were obtained by fitting data to models of competitive, noncompetitive, and uncompetitive inhibition (Segal, 1975) with the Solver function in Microsoft Excel 97 (Microsoft, Redmond, WA).
Immunofluorescent Localization of MRP in Caco-2 Cells.
Washed confluent monolayers of Caco-2 cells grown on Transwells were fixed at room temperature with 3% paraformaldehyde for 15 min to preserve three-dimensional structure. Cells were then permeabilized for 10 min with 0.1% Triton X-100 and blocked for 30 min with 5% normal goat serum. Inserts were incubated for 1 h with antibodies raised against MRP2 (M2I-4, M2III-6) or MRP1 (MRPr1, MRPm6). Primary antibody binding was detected with Alexa 488 goat anti-mouse IgG (MRPm6, M2I-4, M2III-6) or fluorescein isothiocyanate anti-rat IgG (MRPr1; Molecular Probes, Eugene, OR). Cells were incubated for 3 min with propidium iodide for counterstaining of nucleic acids. Alternately, fixed Caco-2 monolayers on polycarbonate membranes were cut from Transwells, embedded in O.C.T. Compound (Miles, Elkhart, IN) and frozen with liquid nitrogen. Frozen sections (5 μm) were cut perpendicular to the membrane. Sections were then permeabilized and stained as described above. Stained monolayers and sections were examined with a Bio-Rad MRC 1024 laser scanning confocal microscope (Hercules, CA). To control for nonspecific binding, matching inserts were treated similarly but with the omission of the primary antibody.
Results
Transcellular Efflux of Quercetin 4′-β-Glucoside.
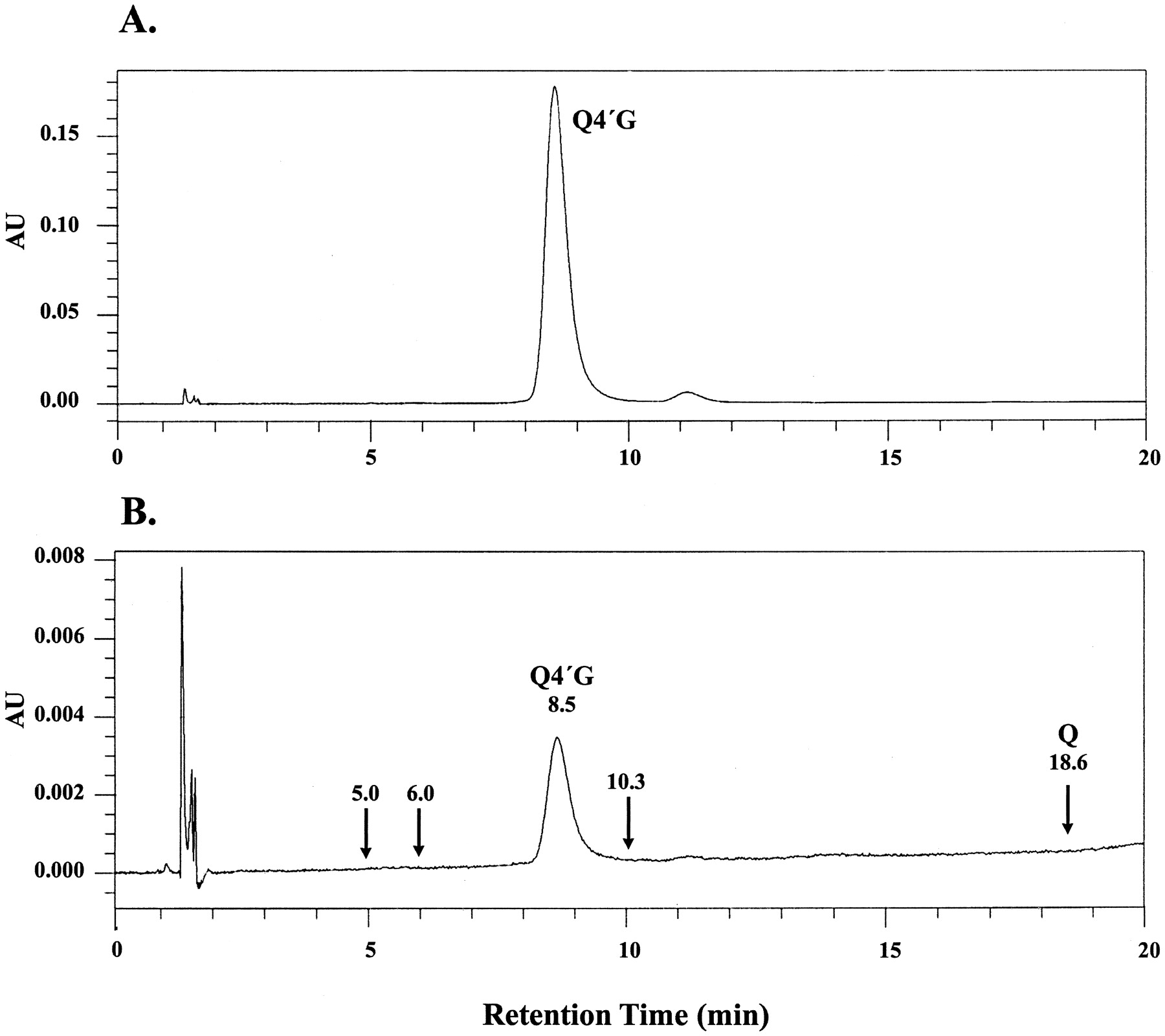
Efflux of quercetin 4′-β-glucoside was monitored by a molecularly specific HPLC method at 370 nm as depicted in Fig. 1. Formation of quercetin or quercetin glucuronides was not observed. The latter was a distinct possibility based on a previous study in the rat intestine (Crespy et al., 1999). However, formation of the glucuronides was ruled out based on the absence of peaks at the retention times (5.0, 6.0, and 10.3 min) identified for the three glucuronides formed when quercetin was incubated with human liver microsomes or recombinant human UGT1A9 and the cofactor UDP-glucuronic acid (Y. Otake and T. Walle, unpublished data).

HPLC of quercetin 4′-β-glucoside in Caco-2 transport experiments. A, loading solution (50 μM). B, apical solution 60 min after loading the basolateral side with quercetin 4′-β-glucoside. The same volume (100 μl) was injected in both tracings. Arrows denote retention times identified for quercetin (Q) and quercetin glucuronides. AU, absorption units; Q4′G, quercetin 4′-β-glucoside.
The efflux of quercetin 4′-β-glucoside across Caco-2 cell monolayers was examined for concentration dependence. Transport of quercetin 4′-β-glucoside was determined in the basolateral-to-apical direction over the concentration range of 10 to 300 μM (Fig.2A). The apparent permeability coefficient decreased significantly with increased concentration over the range examined, from 3.87 ± 0.40 × 10−6 cm s−1 at 10 μM to 0.75 ± 0.06 × 10−6 cm s−1 at 300 μM, consistent with a saturable secretory mechanism. The solubility of quercetin 4′-β-glucoside was determined to be >300 μM and, thus, did not influence the results shown in Fig. 2A.
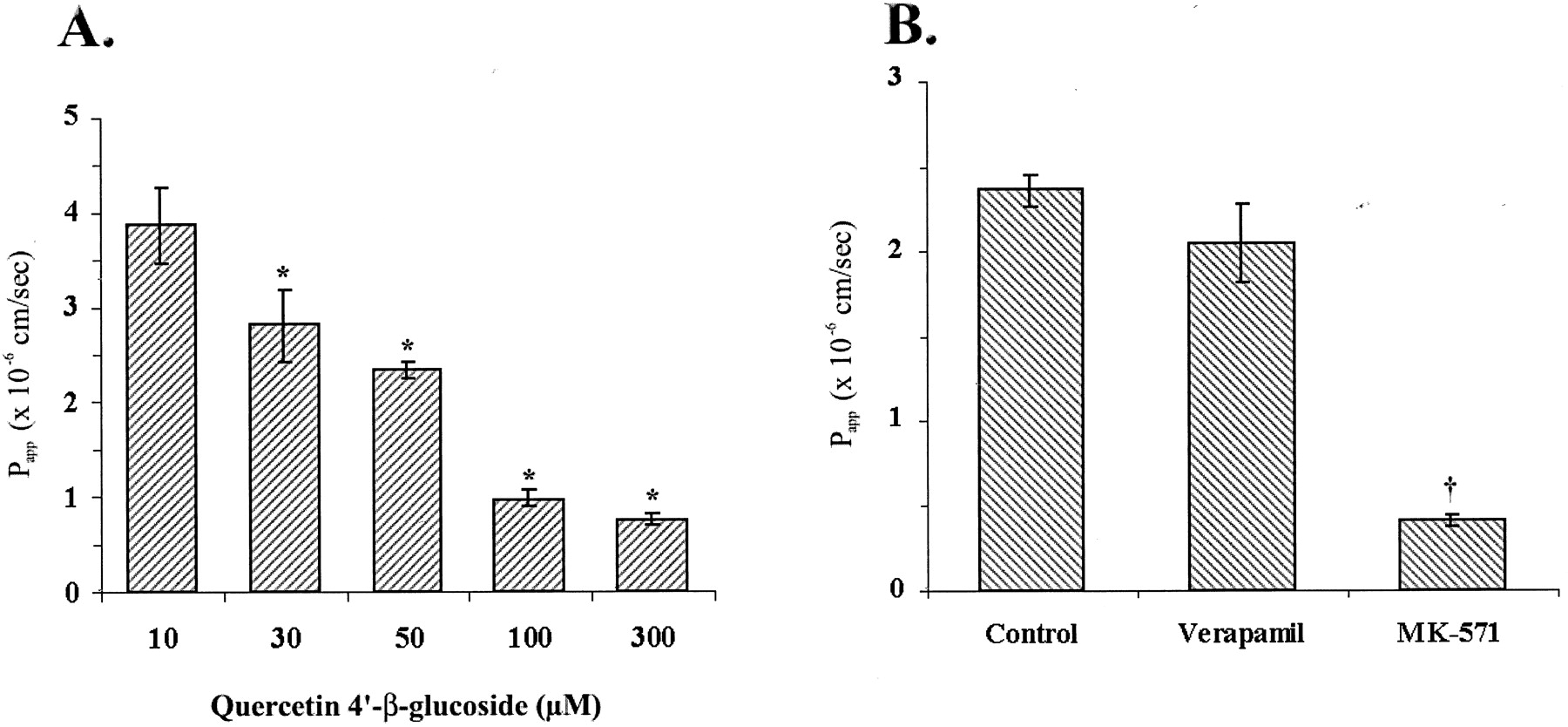
A, concentration-dependent efflux of quercetin 4′-β-glucoside. Washed Caco-2 cell monolayers were loaded on the basolateral side with quercetin 4′-β-glucoside. Samples were collected for analysis after 1 h. Data represent the mean ± S.E. of 4 to 17 independent observations at each concentration. *P < .05, significantly lower than at 10 μM. B, inhibition of quercetin 4′-β-glucoside efflux. Washed Caco-2 cell monolayers were preincubated for 30 min with PBS (control) or with 50 μM verapamil or 50 μM MK-571 in PBS. Buffer ± inhibitor was then replaced and 50 μM quercetin 4′-β-glucoside ± inhibitors was loaded on the basolateral side. Samples were collected for analysis after 1 h. Data represent the mean ± S.E. of five independent observations. †P < .0001, significantly less than control experiments.
To test the hypothesis that a transporter is responsible for the efflux and lack of absorption of quercetin 4′-β-glucoside, we examined the role of P-glycoprotein and the MRPs by measuring efflux in the presence of selective inhibitors (Fig. 2B). Verapamil, an inhibitor of P-glycoprotein, did not significantly alter the flux of quercetin 4′-β-glucoside in the basolateral-to-apical direction,Papp of 2.36 ± 0.09 × 10−6 cm s−1 in control inserts versus 2.04 ± 0.23 × 10−6 cm s−1 with 50 μM verapamil. The presence of 50 μM MK-571, an MRP inhibitor, resulted in a greater than 80% reduction in apparent permeability, 0.40 ± 0.04 × 10−6 cm s−1(P < .001). This finding in combination with the previous detection in our laboratory of MRP2 but not MRP1 in Caco-2 cells (Walle et al., 1999b) implicated a role for MRP2 in the transcellular efflux of quercetin 4′-β-glucoside.
Kinetic Properties of Quercetin 4′-β-Glucoside Efflux
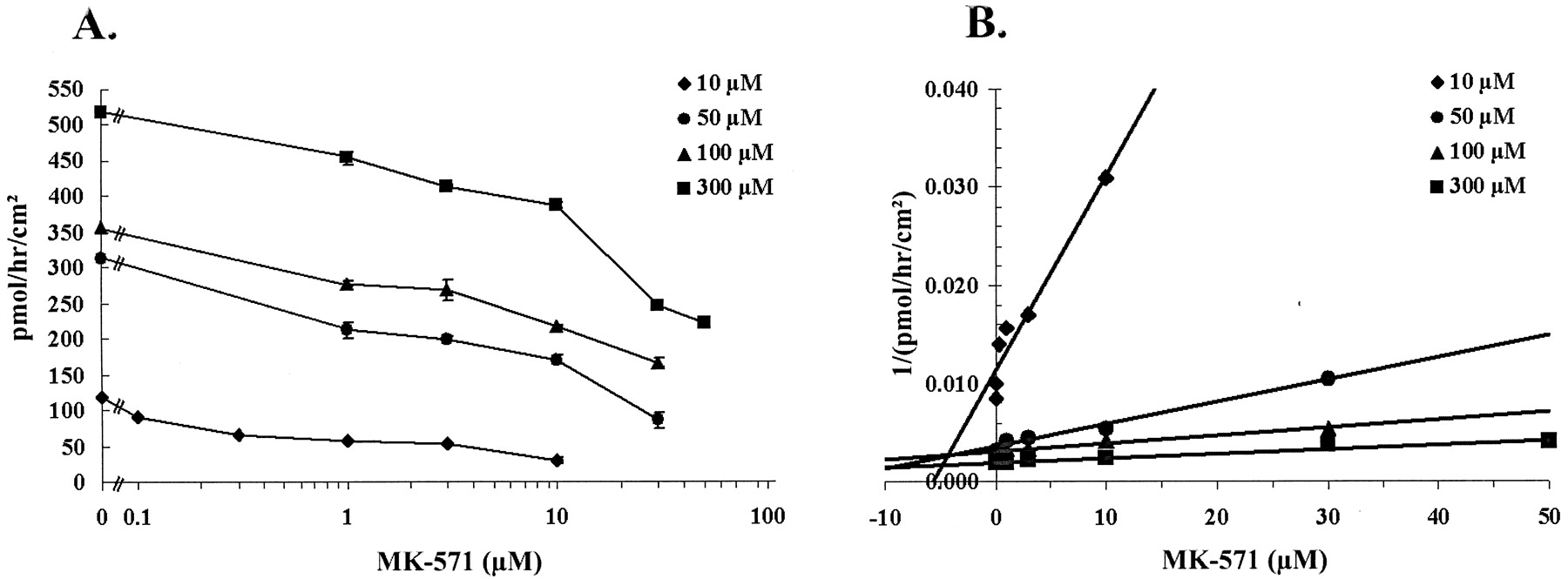
To examine the mechanism of inhibition and determine apparent kinetic constants, we examined quercetin 4′-β-glucoside efflux over a range of substrate and MK-571 concentrations, 10 to 300 μM and 0.1 to 50 μM, respectively. Data were acquired in three separate experiments each with triplicate determinations at each substrate and inhibitor concentration. Both transport of quercetin 4′-β-glucoside and inhibition of transport by MK-571 demonstrated concentration dependence (Fig.3). Data were fitted to one- and two-component models of competitive, noncompetitive, and uncompetitive inhibition with the Solver function in Microsoft Excel 97. Results obtained with a two-component model were not different from those obtained with a one-component model, and the simpler model was adopted. The best fit was obtained with a one-component model of competitive inhibition. Data are summarized in Table1.

Concentration-dependent inhibition of quercetin 4′-β-glucoside transport by MK-571. Washed Caco-2 cell monolayers were preincubated for 30 min with MK-571 in PBS. Buffer ± inhibitor was then replaced and quercetin 4′-β-glucoside ± inhibitor was loaded on the basolateral side. Samples were collected for analysis after 1 h. A, data represent the mean flux ± S.E. of triplicates from three independent experiments. B, Dixon plot of data presented in A.
Apparent kinetic constants derived from the concentration dependent inhibition of quercetin 4′-β-glucoside transport by MK-571
Transcellular Absorption of Quercetin 4′-β-Glucoside
In our previous studies, we have demonstrated a lack of apical to basolateral absorption of quercetin 4′-β-glucoside across Caco-2 monolayers. Efflux of the monoglucoside across the apical membrane by MRP2 is one potential explanation for this observation. To test this possibility we examined the apical-to-basolateral flux of quercetin 4′-β-glucoside in the presence of 50 μM MK-571. Transport of quercetin 4′-β-glucoside was not observed at substrate concentrations less than 250 μM. A modest but statistically significant (P < .001) absorption, Papp = 0.032 ± 0.008 × 10−6 cm s−1, was observed at 250 μM quercetin 4′-β-glucoside in the presence of 50 μM MK-571. No absorption was detected in the presence of 50 μM verapamil.
Immunofluorescent Localization of MRP2 in Caco-2 Cell Monolayers.
The subcellular distribution of MRP isoforms was visualized in confluent Caco-2 cell monolayers grown on Transwells by indirect immunofluorescent localization with a laser scanning confocal microscope. Cells were seeded onto polycarbonate inserts and grown under conditions identical to those used to grow cells for transport studies and monolayer integrity was determined by TEER. Consistent with our previous data from Western blot analysis (Walle et al., 1999b), monolayers stained with the anti-MRP1 antibody MRPm6 were void of green signal and appeared similar to control samples where primary antibodies were omitted (Fig. 4, A and B). Identical results were obtained with MRPr1 (data not shown). In contrast, samples stained with the anti-MRP2 antibody M2III-6 demonstrated significant green signal (Fig. 4C) and similar results were observed with M2I-4 (data not shown). Optical sectioning of thez-series perpendicular to the plane of the cell monolayer demonstrated that the subcellular localization of MRP2 was restricted to the apical side of the cells (Fig. 4C, bottom), whereas no evidence for MRP1 was observed (Fig. 4B, bottom). To enhance resolution and verify the apical staining of MRP2, we examined MRP2 localization in 5-μm sections cut perpendicular to the plane of the cell monolayer. Sections stained with M2III-6 showed that MRP2 immunostaining was primarily confined to the apical plasma membrane, with some diffuse intracellular staining (Fig. 4D).

Immunofluorescent localization of MRP2 in Caco-2 cell monolayers. Expression and localization of MRP1 and MRP2 was examined by indirect immunofluorescence (green signal). Nuclei were detected with propidium iodide (red signal). Scale bar, 10 μm. A and the top of B and C are top-down views of the monolayer (original magnification, 40× water). The bottom of B and C are optical sections perpendicular to the plane of the cell layer. A, confluent Caco-2 monolayer in which primary antibodieswere omitted from staining procedure. B, confluent Caco-2 monolayer stained with anti-MRP1 antibodies, MRPm6. C, confluent Caco-2 monolayer stained with anti-MRP2 antibodies, M2III-6. D, MRP2 localization (M2III-6) in a 5-μm frozen section cut perpendicular to the plane of the cell monolayer (original magnification, 100× oil).
Discussion
A large body of evidence supports a beneficial role for flavonoids in human health. Although the flavonoid content in the human diet is significant, dietary forms of quercetin have demonstrated very poor and variable absorption in rats and humans (Gugler et al., 1975; Ueno et al., 1983; Hollman et al., 1997; Manach et al., 1997). To understand the mechanisms governing the intestinal absorption of flavonoids we have examined transport of quercetin 4′-β-glucoside with human Caco-2 cell monolayers, an accepted model of the human intestinal epithelium. Previously, we have shown that although quercetin 4′-β-glucoside is not absorbed across this intestinal epithelium, it is effluxed (Walgren et al., 1998). This unidirectional flux suggested the possibility that an efflux pump was involved, and in this study, we have presented evidence to support this hypothesis. First, the transport of quercetin 4′-β-glucoside in the basolateral-to-apical direction demonstrated saturation over the concentration range of 10 to 300 μM, consistent with a saturable secretory mechanism. Second, the efflux of quercetin 4′-β-glucoside was competitively inhibited by MK-571, a selective inhibitor of the closely related MRP1 and MRP2 isoforms (Jedlitschky et al., 1994; Leier et al., 1994; Büchler et al., 1996). This observation in combination with the identification of MRP2 but not MRP1 in our Caco-2 cell monolayers supports a role for MRP2 in the efflux of quercetin 4′-β-glucoside across the apical membranes of enterocytes.
Although MRP1 was originally associated with chemotherapeutic resistance, MRP isoforms have since been found to be constitutively expressed in a number of tissues. Within the human intestine, Northern blot analysis demonstrates expression of the message for MRP1, MRP2, MRP3, and MRP5 (Kool et al., 1997). Although the spectrum of substrates is similar or identical for MRP1 and MRP2 (Jedlitschky et al., 1994;Leier et al., 1994; Müller et al., 1994), the cellular localization of these two proteins is different. In hepatocytes, MRP1 is present in lateral membranes, whereas MRP2 is restricted to apical canalicular membranes (Büchler et al., 1996; Roelofsen et al., 1999). Similar to MRP1, MRP3 appears to be expressed in the basolateral membrane of human hepatocytes (König et al., 1999). Currently, little is known about the transport characteristics and cellular localization of MRP5.
Although the cellular localization of the MRP isoforms has been determined within the liver, their localization within the gastrointestinal tract is uncertain. Flens et al. (1996) have previously examined MRP1 expression by immunohistochemistry in normal human tissues. Within the small intestine, epithelial cells demonstrated a supranuclear cytoplasmic staining, but the brush border and goblet cells were negative. Despite a recent claim that Caco-2 cells express MRP1 (Gutmann et al., 1999), previous efforts in our laboratory with Western blot analysis have demonstrated the presence of MRP2 but not MRP1 in Caco-2 cells (Walle et al., 1999b). This apparent discrepancy may result from differences in culture conditions because age and seeding density have been shown to influence expression of MRP1 and MRP2 in cultured hepatocytes (Roelofsen et al., 1999). In this study, we have examined confluent Caco-2 cell monolayers for expression of MRP1 and MRP2 by indirect immunofluorescent localization with laser scanning confocal microscopy. Monolayers were grown in Transwells under identical conditions to those used to grow cells for our transport studies. In agreement with our previous finding, we observed expression of MRP2 but not MRP1. The immunostaining of MRP2 was primarily confined to the apical plasma membrane. This pattern of distribution is consistent with the distribution observed in polarized hepatocytes and similar to that observed in MRP2-transfected Madin-Darby canine kidney cell monolayers (Evers et al., 1998). An apical localization of MRP2 within the intestine would place this transporter in a position to both protect against the absorption of compounds from the lumen and secrete agents into the lumen for removal from the body.
Our kinetic studies with intact cell monolayers revealed that the efflux of quercetin 4′-β-glucoside was competitively inhibited by MK-571 with an apparent KI of 5.6 μM. This value is similar to values observed by Paul et al. (1996)with membrane vesicles from NIH 3T3 cells transfected with MRP1 (KI = 2.5 μM) but less potent than values reported by Jedlitschky et al. (1994) who used membrane vesicles from MRP1-transfected HeLa cells (KI = 0.7 μM). Although comparable values from MRP2 membrane vesicles are not available, it has previously been reported that MK-571 is a slightly less potent inhibitor of MRP2 (Büchler et al., 1996).
Inhibition of quercetin 4′-β-glucoside efflux by MK-571 in combination with the identification of MRP2 expression in the apical membranes of Caco-2 cell monolayers strongly implicates a role for MRP2 in the transcellular efflux of this glucoside. The apparent affinity of quercetin 4′-β-glucoside for MRP2 (Km = 43.6 ± 7.4 μM) does not place this agent among the highest affinity substrates for this transporter. However, the apparent affinity does lie within the predicted lumenal concentration of quercetin 4′-β-glucoside after consumption of a single meal in a typical Western diet (Walgren et al., 1998). This is significant because it allows transport to increase in proportion to increased consumption. A transporter with a lowerKm would already be functioning close to the Vmax and would therefore be unable to compensate for such an increase in concentration.
The spectrum of agents recognized as substrates by MRP1 and MPR2 is similar, consisting primarily of organic anions, and is principally composed of glutathione and glucuronic acid conjugates of lipophilic compounds (König et al., 1999). It is therefore surprising that quercetin 4′-β-glucoside, an uncharged molecule, is a substrate for MRP2. Previously, in a preliminary study, we have shown evidence that another glucoside, genistin, is a substrate for MRP2 (Walle et al., 1999a). The finding that genistin and quercetin 4′-β-glucoside are both substrates for MRP2 suggests that a large new class of agents may be substrates for MRP-mediated transport.
In agreement with the hypothesis that MRP2 is limiting transcellular absorption of quercetin 4′-β-glucoside we have previously demonstrated a lack of apical-to-basolateral absorption of this agent across Caco-2 monolayers (Walgren et al., 1998). To further test this hypothesis we have examined the transcellular absorption of quercetin 4′-β-glucoside, at various substrate concentrations, in the presence of 50 μM MK-571. In contrast to control treatments that did not demonstrate absorption at any substrate concentration, a modest absorption was observed at 250 μM quercetin 4′-β-glucoside in the presence of 50 μM MK-571, supporting this hypothesis but also suggesting that an additional transport mechanism may be involved, preventing quercetin 4′-β-glucoside from crossing the basolateral membrane. Toxicity of MK-571 at higher concentrations and a limited solubility range for quercetin 4′-β-glucoside have limited further testing (data not shown).
In summary, this study demonstrates that MRP2 is localized to the apical membrane of Caco-2 cells and that this protein both limits the absorption of and mediates the efflux of quercetin 4′-β-glucoside across Caco-2 cell monolayers.
Acknowledgment
We thank Martina Sedmerova for assistance with the immunofluorescent localization of MRP in Caco-2 cells.
Footnotes
-
Send reprint requests to: Thomas Walle, Ph.D., Medical University of South Carolina, Department of Cell and Molecular Pharmacology and Experimental Therapeutics, 173 Ashley Ave., P.O. Box 250505, Charleston, SC 29425. E-mail:wallet{at}musc.edu
-
↵1 This study was supported by National Institutes of Health Grant GM55561 and Department of Defense, Army Breast Cancer Program Grant DAMD17-98-1-8125.
- Abbreviations:
- MRP
- multidrug resistance-associated protein
- MEM
- minimum essential medium
- TEER
- transepithelial electrical resistance
- Received March 6, 2000.
- Accepted May 5, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}