


Abstract
The primary aim of the present study was to test the hypothesis that amino acid transport systems are involved in absorptive transport of dicysteinylmercury (cysteine-Hg-cysteine). Luminal disappearance flux [JD, fmol min−1 (mm tubular length)−1] of inorganic mercury (Hg2+), in the form of dicysteinylmercury, was measured in isolated perfused S2 segments with various amino acids or amino acid analogs in the luminal compartment under one of two conditions, in the presence or absence of Na+. The control perfusion fluid contained 20 μM dicysteinylmercury. Replacing Na+ in both the bathing and perfusing solutions withN-methyl-d-glucamine reduced theJD of Hg2+ by about 40%. Nine amino acids and two amino acid analogs were coperfused individually (at millimolar concentrations) with dicysteinylmercury. The amino acids and amino acid analogs that had the greatest effect on theJD of Hg2+ werel-cystine, l-serine, l-histidine,l-tryptophan, and 2-(−)-endoamino-bicycloheptane-2-carboxylic acid. The greatest reduction (76%) in the total JD of Hg2+ occurred when l-cystine was coperfused with dicysteinylmercury in the presence of Na+. Overall, the current findings indicate that Hg2+ is transported from the lumen into proximal tubular epithelial cells via amino acid transporters that recognize dicysteinylmercury. In addition, the data indicate that multiple amino acid transporters are involved in the luminal uptake of dicysteinylmercury, including the Na+-dependent low-affinity l-cystine, B0, and ASC systems and the Na+-independent L-system. Furthermore, the transport data obtained whenl-cystine was added to the luminal fluid indicate strongly that dicysteinylmercury is likely transported as a molecular homolog ofl-cystine.
Recent findings from whole animal (Zalups and Barfuss, 1996), luminal membrane-vesicle (Zalups and Lash, 1997), and isolated-perfused-tubule experiments (Cannon et all, 2000) indicate that the primary luminal mechanism by which inorganic mercuric ions gain access to the cytoplasm of proximal tubular epithelial cells is by being transported as a mercuric conjugate of l-cysteine. Based on the bonding characteristics of mercuric ions (Hughes, 1957; Rabenstein, 1989;Ballatori, 1991; Zalups and Lash, 1994) and the thermodynamic stability of mercuric conjugates of sulfhydryl-containing molecules in aqueous solution (Rabenstein, 1989), it has been postulated that the primary mercuric conjugate of l-cysteine that is transported at the luminal plasma membrane is dicysteinylmercury (cysteine-Hg-cysteine). Since the mercuric conjugate dicysteinylmercury is structurally similar to the amino acid l-cystine (cysteine-cysteine), we had hypothesized previously that one or more of the amino acid transport systems that transports l-cystine also transports dicysteinylmercury. Recent findings from isolated perfused tubule experiments of Cannon et al. (2000) strongly support this hypothesis.
Since the majority of amino acid transport systems are not highly specific for any one particular amino acid, it is reasonable to postulate that mercuric conjugates of l-cysteine may also be transported by amino acid transport mechanisms that do not transportl-cystine. These include both Na+-dependent and Na+-independent transport systems. Some of the better described Na+-dependent transporters include system-A, system-ASC, system-B0, and system-B0,+. System A is found in various organs and is involved in the transport of most dipolar amino acids (Hammerman and Sacktor, 1977). Amino acids or amino acid analogs transported by system-A include l-glycine, l-proline, and α-methylamino-isobutyric acid (MeAIB) (Mircheff et al., 1982; Tate et al., 1989). Consequently, these compounds can be used as competitive inhibitors of this transporter. System-ASC is also found in various organs and is known to transport l-serine,l-alanine, and l-cysteine, all of which can serve as effective competitive inhibitors. In addition, system-ASC has been implicated in the transport of cationic or protonated anionic amino acids (Hammerman and Sacktor, 1977; Kragh-Hansen and Sheikh, 1984). System-B0 is similar to system-ASC in its location and transport specificity, except it has a higher affinity for the larger neutral amino acids than system-ASC. System-B0,+, which has been localized in oocytes, blastocysts, and the luminal plasma membrane of enterocytes and renal proximal tubular epithelial cells, is involved in the transport of a wide range of amino acids, including the amino acids,l-alanine, l-valine, l-tryptophan, and l-lysine (Boerner et al., 1986).
There are at least two primary Na+-independent amino acid transport systems reported to be present in the basolateral (Pineda et al., 1999) and luminal membranes of the proximal tubule, respectively. These are system-L and system-b0,+. System-L is located in a variety of organ systems and is involved in the transport of amino acids with large bulky hydrophobic side chains. Examples include l-phenylalanine, l-tryptophan, and the bicyclic amino acid analog 2-(−)-endoamino-bicycloheptane-2-carboxylic acid (BCH) (Hammerman and Sacktor, 1977; Deves and Boyd, 1998). System-b0,+also transports a wide variety of amino acids. Although system-b0,+ is similar to system-B0,+, it appears to discriminate against amino acids having less extensively branched R-groups attached to the β-carbon. This system has a high affinity for neutral and cationic amino acids, and has been shown to transport l-lysine andl-cystine (Boerner et al., 1986).
The primary objective of the present study was to test the hypothesis that known amino acid transport mechanisms are involved in the absorptive transport of dicysteinylmercury in pars recta (S2) segments of the rabbit proximal tubule. The strategy used in this study was to coperfuse isolated S2 segments through the lumen with dicysteinylmercury in the presence or absence of significantly higher concentrations of various amino acids or analogs that are transported by Na+-dependent and Na+-independent amino acid transport systems. This allowed us to determine whether these compounds inhibit the luminal disappearance flux (JD) of Hg2+ in the form of dicysteinylmercury. It was assumed that significant reductions in theJD of Hg2+induced by the presence of an amino acid or amino acid analog would provide evidence implicating a role of specific amino acid transport systems in the lumen-to-cell and/or cell-to-bath transport of dicysteinylmercury in proximal tubular epithelial cells.
Materials and Methods
Hypothesis and Experimental Design
The primary hypothesis tested in the current study is that the mercuric conjugate dicysteinylmercury is transported across the luminal membrane into the epithelial cells lining S2segments of the rabbit proximal tubule via one or more amino acid transport systems. To test this hypothesis, S2segments of the rabbit proximal tubule were perfused with a modified Ringer's solution [artificial perfusion medium (APM)] containing a 4:1 ratio of l-cysteine to Hg2+ to ensure the formation of the mercuric conjugate dicysteinylmercury (at a concentration of 20 μM). The luminal disappearance flux (JD) of Hg2+, presumably in the form of dicysteinylmercury, was measured with and without purported inhibitory substrates of various amino acid transporters in the perfusing medium. To ensure that the potential inhibitor (amino acid or amino acid analog) was at a great enough concentration to compete with dicysteinylmercury for the transport site, a 50 to 250:1 ratio of inhibitor to dicysteinylmercury was maintained. Perfusion was carried out under two conditions, in the presence or complete absence of Na+ in the perfusing and bathing solutions. This allowed us to assess the role of Na+-dependent versus Na+-independent transporters in the uptake of the putative substrate dicysteinylmercury.
Of necessity, we assumed that a decrease inJD of Hg2+ from the luminal fluid that occurred in the presence an amino acid or amino acid analog in the luminal fluid was the result of competitive inhibition. It is not possible to perfuse segments of the proximal tubule with a great enough range of concentrations of dicysteinylmercury to establish a change ofJmax orKm, which is required to delineate between competitive and noncompetitive inhibition. This limitation is the result of the low specific activity of203Hg2+ and the fact that higher concentrations (above 100 μM) of dicysteinylmercury induce toxic effects in the perfused tubule. Consequently, we have assumed that any inhibition is competitive unless other investigators have reported noncompetitive inhibition by the particular amino acid or amino acid analog, as is the case for l-lysine.
Note: Notwithstanding the fact that mercuric ions can bind to various nucleophilic groups, the affinity constant for bonding to thiolate anions is on the order of 1015 to 1020, whereas the affinity constants for bonding to oxygen- or nitrogen-containing ligands, such as carbonyl groups or amino groups, are about 10 orders of magnitude lower (Hughes, 1957). When mercuric ions and at least a 2-fold greater concentration of small thiol-containing molecules are present in aqueous solution, one can predict with high probability [based on the 13C NMR findings of Rabenstein (1989)] that each mercuric ion will form a thermodynamically stable linear II coordinate covalent bond with the thiol group of two molecules of the respective thiol-containing compound. Thus, addition of a 4-fold higher concentration of cysteine (relative to the concentration of inorganic mercury) ensures the formation of thermodynamically stable linear II coordinate covalent complexes between each mercuric ion and two molecules of cysteine. Moreover, the bonding characteristics of mercuric ions (Hughes, 1957;Rabenstein, 1989) predict that they would remain bonded to the thiol group of cysteine, even in the presence of low millimolar concentrations of other nonthiol-containing amino acids or amino acid analogs, which provide a substantial pool of nucleophilic functional groups for mercuric ions to bind.
Assessment of Role of Na+-Dependent Amino Acid Transporters
System-A.
To determine whether system-A is capable of transporting dicysteinylmercury into proximal tubular cells across the luminal membrane, 3 mM l-proline or l-MeAIB were coperfused individually with 20 μM Hg2+and 80 μM l-cysteine.
System-ASC and B0.
The potential role of system-ASC and/or system-B0 in the luminal uptake of dicysteinylmercury was assessed in experiments where 3 mMl-serine was coperfused with 20 μM Hg2+ and 80 μM l-cysteine
System-B0,+.
Proximal tubular segments were perfused with 3 mM l-tryptophan, l-lysine, orl-valine with 20 μM Hg2+ and 80 μM l-cysteine to determine whether system-B0,+ is capable of transporting dicysteinylmercury.
Cystine Transporters.
Assessment of a specific putative, low-affinity Na+-dependent “l-cystine” amino acid transport system in the luminal disappearance of dicysteinylmercury was done by coperfusing 1 mMl-cystine with 20 μM Hg2+ and 80 μM l-cysteine.
Effect of d-Enantiomer of Cystine.
To determine whether the luminal transport of dicysteinylmercury is stereospecific, S2 segments were perfused through the lumen with 20 μM Hg2+, 80 μM l-cysteine, and 1 mM d-cystine. Additional control experiments were carried out in tubules perfused through the lumen with 20 μM Hg2+, 80 μM l-cysteine with or without 1 mM l-cystine. These experiments were carried at the end of all the other transport experiments. Consequently, additional control data were needed to maintain internal consistency.
Assessment of Role of Na+-Independent Amino Acid Transporters
In these experiments, Na+ in both perfusing and bathing solutions was replaced withN-methyl-d-glucamine.
System-L.
The potential role of system-L in the luminal disappearance flux of dicysteinylmercury was assessed by coperfusing 20 μM Hg2+ and 80 μM l-cysteine with 3 mM l-histidine, 3 mM l-tryptophan, 3 mM cycloleucine, 5 mM l-phenylalanine, or 5 mM BCH.
System-b0,+.
Coperfusion of 20 μM Hg2+ and 80 μM l-cysteine with either 3 mM l-lysine or l-cystine was carried out to assess the potential role of system-b0,+in transporting dicysteinylmercury from the luminal fluid into the epithelial cells lining the pars recta of the proximal tubule.
Animals.
Female, New Zealand White, specific pathogen-free rabbits (Myrtle's Rabbitry, Inc. Farm, Thompson Station, TN) were used in the present study. Prior to experimentation, the rabbits were maintained on regular rabbit chow and given water ad libitum. All experiments were conducted according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Composition of Perfusing and Bathing Solutions.
In all experiments, the perfusing and bathing solutions consisted of simple electrolyte solutions. The perfusing solution (APM) contained the following: 140 mM Na+, 140 mM Cl−, 5 mM K+, 2.5 mM Ca2+, 1.2 mM Mg2+, 1.2 mM SO42−, 2 mM HPO42−/H2PO4−, 1 mM d-glucose, and 0.5 mM glutamine. pH was adjusted to 7.4 with 1 M NaOH. To evaluate cytotoxicity of inorganic mercury, we placed the vital dye FD&C green No.3 (809 Da) in the perfusate at a concentration of 250 nM. Final osmolality was adjusted to 290 mOsmol kg−1 H2O, with doubly distilled and deionized water.l-[3H]glucose (50 mCi ml−1, 58.8 mCi mg−1) (34 μM) was used as a volume marker in all experiments and was added to the perfusing solution only. The concentration of Hg2+ (see note below) in the perfusing solution was 20 μM in all experiments. All perfusing solutions containing Hg2+ also contained radioactive mercuric ions (203Hg2+, 33.6 mCi mg−1).
When transport was studied in the presence of Na+, the control perfusing solution was the artificial perfusing medium to which l-proline (3.0 mM),l-valine (3.0 mM), MeAIB (5.0 mM), BCH (5 mM),l-serine (3 mM), l-tryptophan (3 mM),l-phenylalanine (5.0 mM), or l-lysine (3.0 mM) was added. In all experiments, the bathing solution was the Na+-containing APM without the vital dye added.
In experiments where transport was studied under Na+-independent conditions, Na+ in both the perfusing and bathing solutions was replaced with corresponding amounts ofN-methyl-d-glucamine.
Dissection Solution.
The tubular dissection solution was a sucrose/phosphate buffer: 125 mM sucrose, 13.3 mM anhydrous monosodium dihydrogen phosphate, and 56 mM anhydrous disodium monohydrogen phosphate. The pH was adjusted to 7.4 with NaOH or HCl. The osmolarity was adjusted to 290 mOsm/kg of water by adding water or NaCl.
Chemicals.
All other chemicals were obtained from Sigma (St. Louis, MO), unless otherwise noted. The isotope203Hg2+, in the form of mercuric chloride, was obtained from Buffalo Materials Corporation (Buffalo, NY). l-[3H]Glucose (14.6 Ci mmol−1, 1 mCi ml−1) was obtained from PerkinElmer Instruments (Shelton, CT).
Obtaining, Identifying, and Perfusing S2 Segments of Proximal Tubule.
The methods used for obtaining, identifying, and perfusing each tubule on the day of experimentation were the same as those we have described previously (Zalups et al., 1991; Zalups and Barfuss, 1996).
Collecting Samples.
To measure theJD [fmol min−1 (mm tubule length)−1] of Hg2+ from the luminal compartment, three samples (collectates) of luminal fluid exiting a perfused tubular segment were collected from each perfused tubule. The time required to fill the constant volume pipette (≈60 nl) was used to calculate the collection rate (nl min−1). Each collectate sample was added to 8 ml of scintillation fluid (Opti-Fluor; Packard, Meriden, CT). To measure the rate of lumen-to-bath leak [fmol min−1 (mm tubule length)−1] of the volume markerl-[3H]glucose, the aspirated bathing solution from the flow-through bath (0.3 ml) was collected at the rate of 0.25 ml min−1 into 20-ml scintillation vials at 5-min intervals. To each vial, 8 ml of scintillation fluid (Opti-Fluor; Packard) was added. The collectate and bathing fluid samples were then counted in a Beckman 5800 scintillation counter to quantify of the amount 3H and203Hg present in each sample using standard isotopic methods.
Assessment of Cellular and Tubular Pathology.
During each experiment, the perfused tubule was observed microscopically during the entire perfusion process to detect any pathology. Typical pathological changes detected in S2 segments of the proximal tubule exposed to inorganic mercury include cellular swelling, cytoplasmic vacuolization, shedding of brush-border membrane (blebbing) of the apical plasma membrane, and cellular uptake of the vital dye FD&C green.
Calculations.
The calculations used to determine rates of luminal disappearance flux (JD) of Hg2+ and lumen-to-bath leak of the volume maker (l-[3H]glucose) are the same as those described previously (Zalups and Barfuss, 1996).
Statistical Analysis.
A minimum of four tubules was perfused under each experimental condition. Moreover, data for each parameter assessed were obtained from tubular segments isolated from at least two animals. In each perfused tubule, three or more measurements of theJD of Hg2+ were averaged. The mean values for JD from each tubule were used to compute the overall mean and standard error under each experimental condition. Data were first subjected to the Kolmogorov-Smirnov test for normality and then the Levene's test of homogeneity of variance. If both tests were not statistically significant (at P < 0.05) a one-way analysis of variance and Tukey's honestly significant difference post hoc test was performed with a significance level set at P< 0.05. If a set of data failed the normality test or the test for homogeneity of variance, the nonparametric Kruskal-Wallis analysis of variance by ranks, followed by a Mann-Whitney U test analysis was performed with the level of significance set atP < 0.05.
Results
Effect of Various Amino Acids on Na+-Dependent Transport of Hg2+
Control data for the experiments designed to assess inhibition of the JD of dicysteinylmercury under Na+-dependent conditions were obtained from tubular segments perfused with the APM containing 20 μM Hg2+ and 80 μMl-cysteine. During 30 min of perfusion, theJD of Hg2+averaged approximately 102 fmol min−1 (mm tubular length)−1(Fig.1).

Luminal disappearance flux (JD) of Hg2+in isolated S2 segments of the rabbit proximal tubule perfused with 20 μM Hg2+ and 80 μM l-cysteine, with or without 3.0 mM l-proline, 3.0 mM l-valine, or 5.0 mM MeAIB in the perfusing solution. All tubules were perfused and bathed with solutions containing 140 mM Na+. Each column represents the mean ± S.E. obtained from at least four perfused tubular S2 segments. ∗, significantly different (P < 0.05) from the mean value obtained from the control tubules perfused with 20 μM Hg2+ and 80 μM cysteine without any additional amino acids or amino acid analogs in the perfusing solution.
To assess whether amino acids transported by systems-A and/or B0,+ are involved in the luminal transport of dicysteinylmercury, 3 mM l-proline, 3 mMl-valine, or 5 mM MeAIB was added to the control perfusate. The presence of each of these amino acids in the perfusate did not have a significant effect on the JD of Hg2+(Fig. 1). Additionally,l-proline and l-valine had no effect on the concentration of Hg2+ in the collectate, while the presence of MeAIB in the perfusate caused a 250% increase in the concentration of Hg2+ in the collectate (Table 1).
Concentration of Hg2+ in collectate, leak rate ofl-[3H]glucose, and visual toxicity data from S2 segments of the rabbit proximal tubule perfused with 20 μM Cys-Hg-Cys (20 μM Hg2+ and 80 μM l-cysteine) alone (control) or with various potential competitive inhibitors in the presence of 140 mM Na1-213
The mean JD of Hg2+ (in the form of dicysteinylmercury) was reduced markedly when either 3 mM l-lysine or 3 mM l-serine was present in the control perfusion solution (Fig. 2). More specifically, addition of 3 mM l-lysine or 3 mMl-serine to the perfusate caused an approximate 52 or 53% decrease in the JD of Hg2+, respectively. The concentration of Hg2+ in the samples of collectate increased by 290 and 345%, respectively, when l-lysine orl-serine were coperfused with dicysteinylmercury (Table 1). These particular experiments were designed to determine whether the amino acids transported by systems-ASC, B0, B0,+, and/or b0,+ are involved with the luminal transport of dicysteinylmercury.

Luminal disappearance flux (JD) of Hg2+in isolated S2 segments of the rabbit proximal tubule perfused with 20 μM Hg2+ and 80 μM l-cysteine, with or without 3.0 mM l-lysine or 3.0 mM l-serine in the perfusing solution. All tubules were perfused and bathed with solutions containing 140 mM Na+. Each column represents the mean ± S.E. obtained from at least four perfused tubular S2 segments. ∗, significantly different (P < 0.05) from the mean value obtained from the control tubules perfused with 20 μM Hg2+ and 80 μMl-cysteine without any additional amino acids or amino acid analogs in the perfusing solution.
Relative to control values, significant decreases in theJD of Hg2+ were detected in S2 segments perfused through the lumen with 5 mM BCH (38% decrease), 5 mM cycloleucine (57% decrease), 3 mM l-tryptophan (62% decrease), or 5 mMl-phenylalanine (65% decrease) (Fig.3). Among these conditions evaluated, theJD of Hg2+ was lowest in the tubules perfused withl-phenylalanine and was highest in tubules perfused with BCH. The percentage of change in the concentration of Hg2+ in the samples of collectate increased by 240, 330, 360, and 470%, respectively, when BCH, cycloleucine,l-tryptophan, orl-phenylalanine was present in the perfusate (Table 1). The effect of these amino acids on theJD of Hg2+ was examined to determine whether the amino acids transported System-L participates in the absorptive transport of dicysteinylmercury.

Luminal disappearance flux (JD) of Hg2+in isolated S2 segments of the rabbit proximal tubule perfused with 20 μM Hg2+ and 80 μM l-cysteine, with or without 5.0 mM BCH, 5.0 mM cycloleucine, 3.0 mMl-tryptophan, or 3.0 mM l-phenylalanine in the perfusing solution. All tubules were perfused and bathed with solutions containing 140 mM Na+. Each column represents the mean ± S.E. obtained from at least four perfused tubular S2segments. ∗, significantly different (P < 0.05) from the mean value obtained from the control tubules perfused with 20 μM Hg2+ and 80 μM cysteine without any additional amino acids or amino acid analogs in the perfusing solution.
Among the experiments in which both perfusing and bathing solutions contained 140 mM Na+, the greatest effect on the JD of Hg2+ was detected between the group of tubules perfused with 1 mM l-cystine and the group of control tubules (Fig. 4). TheJD of Hg2+ in the tubules perfused with 1 mM l-cystine averaged approximately 25 fmol min−1 (mm tubular length)−1, which is 76% lower than the averageJD of Hg2+ in the control tubules. In addition, the concentration of Hg2+ in the samples of collectate from the tubules perfused with 1 mM l-cystine was 470% greater than that in the samples of collectate from the control tubules (Table 1). These experiments were designed to determine whether similarity in molecular homology between dicysteinylmercury and the amino acid cystine plays an important role in the transport of dicysteinylmercury by transport-proteins involved in proximal tubular absorption of cystine.

Luminal disappearance flux (JD) of Hg2+in isolated S2 segments of the rabbit proximal tubule perfused with 20 μM Hg2+ and 80 μM L-cysteine, with or without 1.0 mM l-cystine in the perfusing solution. All tubules were perfused and bathed with solutions containing 140 mM Na+. Each column represents the mean ± S.E. obtained from at least four perfused tubular S2 segments. ∗, significantly different (P < 0.05) from the mean value obtained from the control tubules perfused with 20 μM Hg2+ and 80 μM l-cysteine without any additional amino acids or amino acid analogs in the perfusing solution.
To determine whether the effect on l-cystine on the luminal uptake of dicysteinylmercury is stereospecific, the effect ofd-cystine on the luminal uptake of dicysteinylmercury was examined. With 140 mM Na+ present in the luminal and basolateral fluid compartments, coperfusing S2 segments with 20 μM Hg2+ and 80 μM l-cysteine with 1 mMd-cystine did not result in a significant reduction in theJD of Hg2+ (in the form of dicysteinylmercury). The transport data obtained from the separate set of control tubules (matched for these experiments) confirmed that 1 mM l-cystine inhibited theJD of dicysteinylmercury to a greater extent than any other amino acid (by 85% in these additional tubules). Thus, two independent series of experiments indicate thatl-cystine is a very effective inhibitor of the luminal uptake of inorganic mercury (in the form of dicysteinylmercury) in proximal tubular segments.
Cellular Toxicity with Na+ Being Present in Both Perfusing and Bathing Solutions.
In all of the aforementioned experiments, no visual evidence of acute cellular toxicity was detected. However, lumen-to-bath leak ofl-[3H]glucose was slightly but significantly greater (2–3 times) in four groups (control, MeAIB, BCH, and l-phenylalanine) than that in normal tubules not perfused with Hg2+ [≈2.5 fmol min−1 (mm tubule length)−1] (Barfuss and Schafer, 1981). By contrast, the presence of l-valine, l-proline,l-lysine, l-serine, cycloleucine,l-tryptophan, or l-cystine in the luminal compartment had no effect on the rate of leak ofl-[3H]glucose (Table 1).
Effect of Various Amino Acids on Na+-Independent Transport of Hg2+
Replacement of Na+ in both the perfusing and bathing solutions withN-methyl-d-glucamine caused a significant reduction (40%) in JD of Hg2+(Fig. 5). The concentration of Hg2+ in the samples of collectate increased by 260%, compared with the concentration of Hg2+ in the samples of collectate from the tubules perfused in the presence of Na+ (control group in Table 1).

Effect of replacing Na+ withN-methyl-d-glucamine, in both bathing and perfusing solutions, on the luminal disappearance flux (JD) of Hg2+in isolated S2 segments of the rabbit proximal tubule perfused with 20 μM Hg2+ and 80 μM l-cysteine. Each column represents the mean ± S.E. obtained from at least four perfused tubular S2 segments. ∗, significantly different (P < 0.05) from the mean value obtained from the control tubules perfused with 20 μM Hg2+ and 80 μMl-cysteine with Na+-containing perfusion and bathing solutions.
Effects of l-Lysine.
After Na+ had been replaced withN-methyl-d-glucamine, in both the perfusing and bathing solutions, no significant difference could be detected in the JD of Hg2+ between S2 segments perfused through the lumen with 3.0 mM l-lysine and the corresponding group of control S2segments (Fig. 6). The concentration of Hg2+ in the samples of collectate from these two groups was also not significantly different. These experiments were preformed to assess the extent the amino acid transport systems L, y+, and b0,+ play in the luminal transport of dicysteinylmercury.
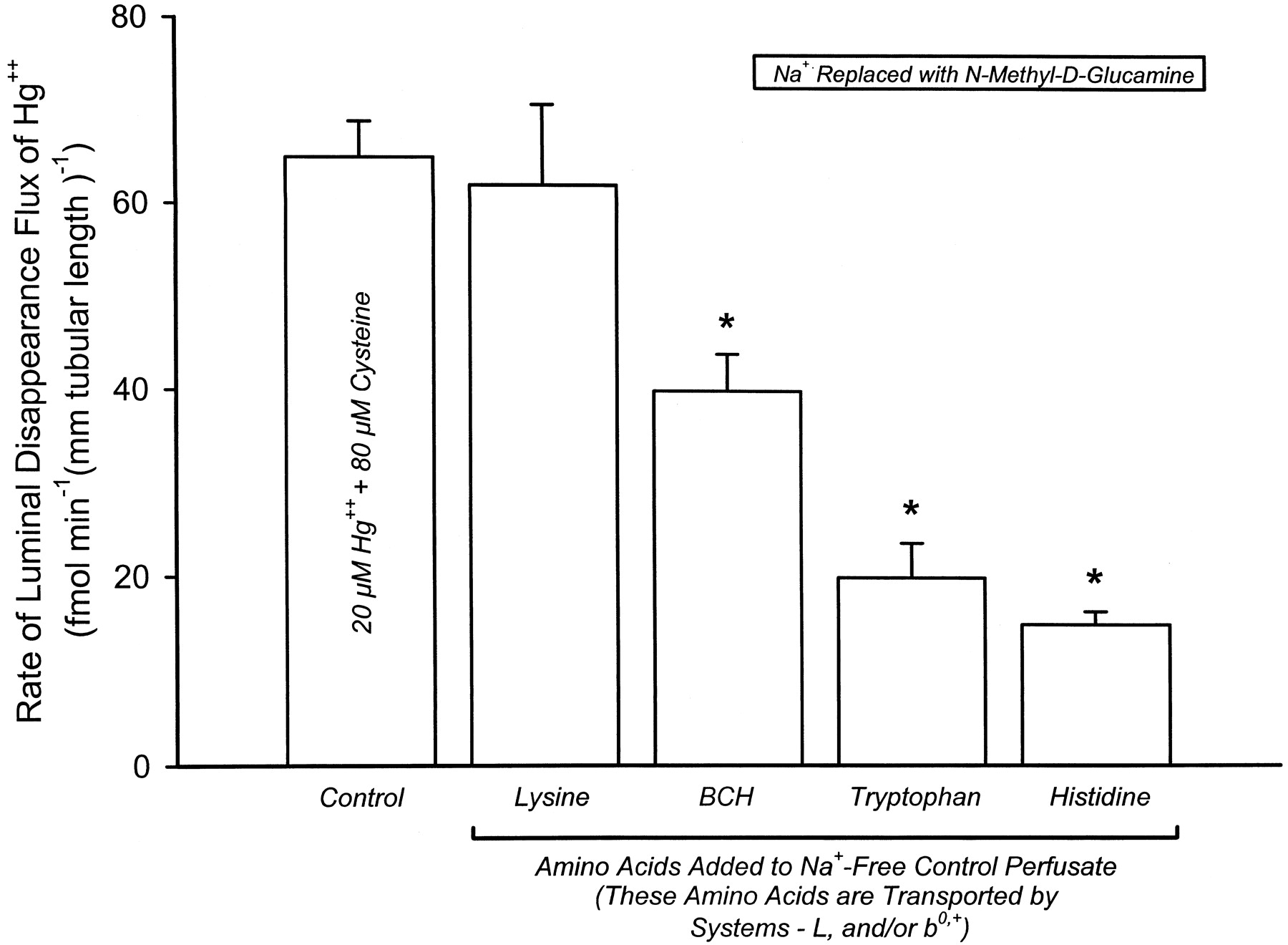
Luminal disappearance flux (JD) of Hg2+ in isolated S2 segments of the rabbit proximal tubule perfused with 20 μM Hg2+ and 80 μM l-cysteine, with or without 3.0 mM lysine, 5.0 mM BCH, 3.0 mM l-tryptophan, or 3.0 mM l-histidine in the perfusing solution under Na+-free conditions. All tubules were perfused and bathed with solutions containing 140 mMN-methyl-d-glucamine. Each column represents the mean ± S.E. obtained from at least four perfused tubular S2 segments. ∗, significantly different (P < 0.05) from the mean value obtained from the control tubules perfused with 20 μM Hg2+ and 80 μMl-cysteine without any additional amino acids or amino acid analogs in the perfusing solution.
In contrast to the lack of effect of l-lysine on theJD of Hg2+, addition of BCH (5 mM), l-tryptophan (3 mM), orl-histidine (3 mM) to a Na+-free perfusate caused additional significant reductions in the JD of Hg2+ (Fig. 6). More precisely, when BCH,l-tryptophan or l-histidine was present in the Na+-free perfusate containing 20 μM Hg2+ and 80 μM l-cysteine, the JD of Hg2+was 38, 68, and 77% lower, respectively, than theJD value obtained in the corresponding control tubules perfused under Na+-free conditions. The concentration of Hg2+ in the samples of collectate increased by 37, 87, or 93%, respectively, when BCH, l-tryptophan, orl-histidine was present in the perfusate (Table2). These experiments were preformed to determine whether the amino acid transport system-L plays a role in the luminal transport of dicysteinylmercury.
Concentration of Hg2+ in collectate, leak rate ofl-[3H]glucose, and visual toxicity data from S2 segments of the rabbit proximal tubule perfused with 20 μM Cys-Hg-Cys (20 μM Hg2+ and 80 μM l-cysteine) alone (control) or with various potential competitive inhibitors in the presence of 140 mM N-methyl-d-glucamine
Cellular Toxicity in Na+-Free Conditions.
Much like when 140 mM Na+ was present in the perfusing and bathing solutions, no visible signs of toxicity were observed during the experiments in which l-lysine, BCH,l-tryptophan, or l-histidine was used as a competitive inhibitor in the presence of 140 mMN-methyl-d-glucamine. There were, however, significant increases in the lumen-to-bath flux of the leak indicator (l-[3H]glucose) with the use of some of the amino acids. The rate of leak was not elevated significantly when l-lysine, BCH, orl-tryptophan was present in the perfusate containing N-methyl-d-glucamine, when compared with that detected in the Na+-free control experiments. Only when l-histidine was present in the perfusate, was the rate of leak ofl-[3H]glucose increased (≈3 times).
Discussion
Several lines of in vivo and in vitro evidence indicate that the luminal absorptive transport of Hg2+ in the renal proximal tubule is linked to the activity of γ-glutamyltransferase and cysteinylglycinase (Berndt et al., 1985;Tanaka et al., 1990; Tanaka-Kagawa et al., 1993; deCeaurriz et al., 1994; Zalups, 1995; Cannon et al., 2000). In the kidney, these enzymes are found almost exclusively in the luminal plasma membrane of proximal tubular epithelial cells. Consequently, the actual luminal uptake of Hg2+ in the proximal tubule appears to involve the transport of some cleavage product(s) formed by the actions of these enzymes. Because the activities of both γ-glutamyltransferase and cysteinylglycinase are very high in the luminal compartment of proximal tubular segments, the most likely primary species of Hg2+ transported at the luminal membrane would appear to be a mercuric conjugate ofl-cysteine (dicysteinylmercury).
Substantive in vivo and in vitro data support the hypothesis that dicysteinylmercury (cysteine-Hg-cysteine) is one of the primary forms of Hg2+ taken up by proximal tubular epithelial cells (Zalups and Barfuss, 1996; Zalups and Lash, 1997). By far, the most convincing evidence supporting the luminal transport of a mercuric conjugate of l-cysteine comes from the isolated perfused tubule studies of Cannon et al. (2000). These investigators demonstrated in isolated perfused S2 proximal tubular segments that the rates of luminal uptake (luminal disappearance flux, JD) of Hg2+ in the form of dicysteinylmercury were approximately 2-fold greater than the rates of luminal uptake of Hg2+ in the form of mercuric conjugates of either glutathione or cysteinylglycine. They also demonstrated that addition of millimolar concentrations of l-lysine or cycloleucine to a perfusate containing 20 μM Hg2+ and 80 μM l-cysteine (the same as in the present study) caused an approximate 50% reduction in the net rate of absorptive transport of Hg2+. Collectively, these findings led the investigators to postulate that luminal uptake of Hg2+, presumably in the form of dicysteinylmercury, occurs through one or more transporters shared byl-cystine and the dibasic amino acidl-lysine.
Consistent with the isolated perfused tubule-findings of Cannon et al. (2000) are the results of the Richardson et al. (1975). These researchers demonstrated that the uptake of Hg2+in renal cortical slices was reduced when the slices were exposed to mercuric conjugates of l-cysteine and an excess of the amino acid l-histidine or l-lysine. In addition, Wei et al. (1999) have demonstrated in isolated fragments of the rabbit proximal tubule that proximal tubular cells may sequester several mercuric conjugates, including mercuric conjugates of cysteine, by a neutral amino acid transport mechanism. It should be kept in mind, however, that uptake in tissue slices and nonperfused isolated tubular fragments probably results primarily via transport mechanisms located on the basolateral membrane. Other than these findings, there has been a paucity of data dealing with potential mechanisms involved in the renal tubular uptake and transport of Hg2+ in the kidney. This deficit served as the primary impetus for the present study. The experiments in the present study were designed specifically to investigate the potential role of some of the better characterized amino acid transport systems in the disappearance of the mercuric conjugate dicysteinylmercury from the luminal fluid of the S2 segment of the rabbit proximal tubule. The Na+-dependent amino acid transporters targeted were systems A, ASC, B0, and B0,+, while the Na+-independent amino acid transporters targeted were systems L and b0,+.
Na+-Dependent Transport of Dicysteinylmercury.
According to our present findings, system-A does not appear to be involved significantly in the luminal uptake of dicysteinylmercury in the proximal tubule. This conclusion is based on the findings that the luminal uptake of 20 μM dicysteinylmercury was not inhibited significantly by millimolar concentrations of l-proline or MeAIB, which are established substrates for the system-A amino acid transporter (Fig. 1) (Hammerman and Sacktor, 1977).
The flux data from the present study also indicate that the Na+-dependent, cationic amino acid transporter, system B0,+, which has been localized in the brush-border membrane of proximal tubular epithelial cells (Boerner et al., 1986), does not play a significant role in the luminal uptake of dicysteinylmercury in pars recta segments of the proximal tubule. Evidence supporting this assertion comes mainly from the experiments in which addition of l-valine to the perfusing medium did not affect significantly the luminal JD of Hg2+ (Fig. 1). It was surprising thatl-valine did not have an effect, whilel-lysine had profound effects, on the luminal absorptive transport of dicysteinylmercury (Fig. 2). This is particularly surprising because both of these amino acids are known to be competitive inhibitors of system-B0,+ (Boerner et al., 1986). These and other data from the present study suggest that the inhibitory effects of l-lysine on theJD of Hg2+ (as dicysteinylmercury) are most likely due to inhibition of system-ASC, rather than system-B0,+. This effect ofl-lysine on the uptake of dicysteinylmercury may be due actually to a noncompetitive inhibitory effect on the “ASC” amino acid transport system. It is thought that the positive charge onl-lysine occupies the Na+-binding site on the “ASC” transporter. This argument has been used to explain the effects ofl-lysine on transport processes in erythrocytes (Young et al., 1988).
It appears that the Na+-dependent luminal transport of dicysteinylmercury most likely involves, at least in part, a neutral amino acid transport system like system-ASC and/or B0. The classical ASC and B0 transporter systems have been implicated in the transport of the amino acid l-serine (Kragh-Hansen and Sheikh, 1984). Support for our hypothesis that system-ASC and/or B0 is involved in the luminal transport of dicysteinylmercury comes from the experiments wherel-serine effectively inhibited approximately 53% of theJD of Hg2+ (as dicysteinylmercury) (Fig. 2). This decrease inJD is also reflected in the large increase in the collectate concentration of Hg2+that resulted from inhibition of uptake of Hg2+at the luminal membrane (Table 1). In addition, BCH (a substrate for the Na+-dependent B0system) inhibited the JD of Hg2+ by about 40 fmol min−1 (mm tubular length)−1 (Fig. 3), while it decreased theJD of Hg2+ by only 20 fmol min−1 (mm tubular length)−1 (Fig. 6) under Na+-free conditions, which implicates system B0 in the transport of dicysteinylmercury.
Dicysteinylmercury may also be transported by a low-affinity transporter that is specific for l-cystine only (Gunther and Silbernagl, 1981). Addition of l-cystine to the perfusate had the most profound effects on theJD of dicysteinylmercury, resulting in a 76% (to 85%) reduction in the rate of uptake (Fig. 4). By marked contrast, addition of the d-enantiomer of cystine to the perfusion medium did not result in an inhibitory effect on the luminal absorptive transport of dicysteinylmercury. These findings indicate strongly that dicysteinylmercury andl-cystine share a common, stereospecific transporter. These findings also support the hypothesis that dicysteinylmercury enters into proximal tubular cells across the luminal plasma membrane as a molecular homolog of cystine.
Na+-Independent Transport of Dicysteinylmercury.
Findings from the current study show clearly that the luminal transport of dicysteinylmercury uses both Na+-dependent and Na+-independent transport mechanisms to absorb this mercuric conjugate from the luminal fluid. Primary support for this conclusion comes from the data showing that an approximate 40% reduction in the JD of Hg2+ occurs in S2 segments when Na+ in both perfusing and bathing solutions is replaced with N-methyl-d-glucamine (Fig. 5).
The Na+-independent cationic amino acid transport system-b0,+ was shown not to be involved significantly in the luminal uptake of dicysteinylmercury in the current study. Interestingly, this system is known to transportl-lysine (Boerner et al., 1986). With the addition of 3 mMl-lysine to the perfusion medium, there was no change in the luminal transport of dicysteinylmercury under Na+-free conditions (Fig. 6). This lack of effect of l-lysine suggests that system-b0,+is not involved in the luminal uptake of dicysteinylmercury.
Data from the present study do, however, indicate strongly that the Na+-independent system-L contributes substantially to the luminal disappearance of dicysteinylmercury in isolated S2 segments of the proximal tubule. It has been demonstrated that system-L transports neutral amino acids and amino acid analogs with bulky hydrophobic side chains, such asl-phenylalanine, cycloleucine, l-tryptophan,l-histidine, and BCH (Hammerman and Sacktor, 1977; Deves and Boyd, 1998). BCH, l-tryptophan,l-phenylalanine, and cycloleucine inhibited individually the luminal disappearance of dicysteinylmercury as indicated by the reduced luminal uptake and increased concentration of Hg2+ in the collectate when Na+ was present in the perfusion and bathing solutions (Fig. 3; Table 1). By inference, cycloleucine is transported by system-L because of the bulky hydrophobic side chain on this molecule. Indeed, l-phenylalanine and cycloleucine have been shown to exhibit mutual inhibition (Gunther and Silbernagl, 1981). BCH and l-tryptophan were also able to reduce significantly the JD of dicysteinylmercury when Na+ was absent from the perfusate (Fig. 6; Table2). Significant reductions in the luminal uptake of dicysteinylmercury also occurred under Na+-dependent conditions when cycloleucine was present in the perfusing solution (Fig. 3). Collectively, these data support the hypothesis that the Na+-independent transport of dicysteinylmercury involves an “L”-type neutral amino acid transporter. The site of this inhibition is probably located at the basolateral membrane for the cell-to-bath exit of dicysteinylmercury, since system-L is thought to be located only at this site (Pineda et al., 1999).
Potential Nonspecific Effects.
One might be led to speculate that some of the decreases in JDdetected with the use of millimolar concentrations of amino acids could be due, in part, to competitive binding of mercuric ions to the nucleophilic functional groups (such as carboxyl, amide, and amine groups) present on the amino acids. However, this is very unlikely based on the tremendously strong and thermodynamically stable bond formed between mercuric ions and the sulfur atom of the thiol group of cysteine. To put some perspective on this issue, the reported affinity (constant) for mercuric ions bonding to a thiolate anion is on the order of 1010 times greater than that for mercury bonding to any other nucleophilic group (Hughes, 1957).
The lack of inhibition in the luminal uptake of dicysteinylmercury whenl-proline, l-valine, or d-cystine was present in the perfusion fluid leads one to conclude that competitive binding to nonthiol-containing nucleophiles is not a major contributing factor responsible for the reductions in the luminal uptake of dicysteinylmercury detected with the use of certain amino acids and analogs. The data obtained with the l- andd-enantiomers of cystine also support this conclusion. In fact, these data support the hypothesis that the luminal disappearance of dicysteinylmercury is due to absorptive transport at the site(s) of stereospecific amino acid transporters.
Another potential nonspecific effect that could potentially affect transport of mercuric conjugates relates to the influence of the removal of Na+ from the perfusing and bathing solutions in the experiments designed to assess Na+-independent transport. Indeed, replacement of Na+ withN-methyl-d-glucamine could have altered intracellular levels of both H+ and Ca2+ via Na+ exchangers (Geigel et al., 1989; Frindt and Windhager, 1990; Lyu et al., 1991), which could have an effect on transport parameters. Only additional investigations can provide substantive information on the potential effects of altered intracellular pH and/or Ca2+on the luminal absorptive transport of dicysteinylmercury in the presence and/or absence of various amino acids. One point is certain, however, and that is, if there were significant alterations in the intracellular milieu induced by the removal of Na+, they were not reflected pathologically, since the morphology of the tubular epithelial cells appeared normal throughout the perfusion process. No evidence of cellular swelling, luminal membrane blebbing, uptake of the vital dye FD&C green, and/or cellular vacuolization was detected in the tubules perfused under Na+-free conditions. Moreover, intercellular leak in the tubules was generally within the normal range (Table 2). Therefore, we interpret any decrease in absorptive transport associated with Na+-free conditions as an indication of a specific Na+-dependent absorptive transport process.
The primary conclusions from this study are that the mercuric conjugate dicysteinylmercury is transported from the luminal fluid of S2 segments of the proximal tubule by both Na+-dependent and Na+-independent amino acid transport systems and that these transport systems recognize dicysteinylmercury as a molecular homolog of l-cystine (cysteine-cysteine) or as an independent molecular structure. Moreover, our data indicate that the transport of dicysteinylmercury is stereospecific. The Na+-dependent transport of dicysteinylmercury appears to involve the specific low-affinity l-cystine transporter, system-B0, and system-ASC. The “L”-type amino acid transporter(s) appear to account for most of the Na+-independent transport of dicysteinylmercury from the luminal fluid.
Footnotes
-
This study was supported by grants from the National Institutes of Environmental Health Sciences (ES 05980 to R.K.Z. and D.W.B. and ES 05157 to R.K.Z.). Vernon Cannon was supported by a graduate student minority supplement to Grant ES05157 awarded by the National Institutes of Environmental Health Sciences.
- Abbreviations:
- MeAIB
- α-methylamino-isobutyric acid
- BCH
- 2-(−)-endoamino-bicycloheptane-2-carboxylic acid
- JD
- luminal disappearance flux, fmol min−1 (mm tubular length)−1
- APM
- artificial perfusion medium
- Received November 20, 2000.
- Accepted April 17, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}