


Abstract
We have investigated allosteric interactions of four closely related strychnine-like substances: Wieland-Gumlich aldehyde (WGA), propargyl Wieland-Gumlich aldehyde, strychnine, and brucine with N-methylscopolamine (NMS) on M3 subtype of muscarinic receptor genetically modified in the second or the third extracellular loop to corresponding loops of M2 subtype (M3o2 and M3o3 chimera). The M3o2 chimeric receptor The exhibited no change in either affinity of strychnine, brucine, and WGA or in cooperativity of brucine or WGA, whereas both parameters for propargyl-WGA changed. In contrast, there was a change in affinity of all tested modulators (except for brucine) and in their cooperativity in the M3o3 chimera. Directions of affinity changes in both chimeras were always toward values of the donor M2 subtype, but changes in cooperativity were variable. Compared with the native M3 receptor, strychnine displayed a slight increase in positive cooperativity and propargyl-WGA a robust decrease in negative cooperativity at M3o2 chimera. Similar changes were found in the M3o3 chimera. Interestingly, cooperativity of brucine and WGA at the M3o3 chimera changed from negative to positive. This is the first evidence of constitution of positive cooperativity of WGA by switching sequences of two parental receptors, both exhibiting negative cooperativity. Gradual replacement of individual amino acids revealed that only three residues (NVT of the o3 loop of the M2 receptor) are involved in this effect. Data suggest that these amino acids are essential for propagation of a conformation change resulting in positive cooperativity induced by these modulators.
Muscarinic receptors belong to the superfamily of seven transmembrane spanning segment, G protein-coupled receptors (Fredriksson et al., 2003). Five subtypes of muscarinic receptors that share a high degree of homology in transmembrane segments while connecting extracellular loops displaying more diversity have been identified (Bonner et al., 1987, 1988; Peralta et al., 1987; Bonner, 1989a,b). Selectivity of individual subtypes for classical ligands and particularly for agonists is rather low because the orthosteric binding site is formed by highly homologous transmembrane segments (Lu et al., 2002; Hulme et al., 2003). Besides the classical binding site, muscarinic receptors possess one or more allosteric binding sites (Ellis and Seidenberg, 1989; Ellis et al., 1993; Lazareno et al., 2000; Christopoulos and Kenakin, 2002) whose binding determinants are located on less conserved extracellular loops. Binding of allosteric ligands can induce conformational changes of the receptor that result in changes in receptor affinity for orthosteric agonists and antagonists (Tuček et al., 1990, 1998; Proška and Tuček, 1994; Dong et al., 1995; Jakubík et al., 1995, 1997; Lazareno and Birdsall, 1995). These effects exhibit remarkable receptor subtype selectivity that is also reflected in the functional outcome of receptor stimulation by orthosteric agonists (Lazareno and Birdsall, 1995; Doležal and Tuček, 1998; Birdsall et al., 1999; Lazareno et al., 2004). Allosteric interactions are usually more pronounced at M2 and M4 than at M1, M3, or M5 subtypes of muscarinic receptors (Christopoulos et al., 1999; Christopoulos and Kenakin, 2002), and at least in the case of the allosteric modulator gallamine this difference between M2 and M3 subtypes is not due to preferential coupling with G protein (Tränkle et al., 2001), which is largely determined by the third intracellular loop. Allosteric modulators are thus useful for identifying muscarinic receptor subtype that mediate specific physiological functions in native systems and also offer a great promise for developing strategies of subtype-selective therapeutic interventions.
Allosteric modulator-induced effects on orthosteric ligand affinity is characterized by a factor of cooperativity (α) that denotes direction and maximal change in binding affinity of the orthosteric ligand. The magnitude of α is dependent on the pair of allosteric/orthosteric ligands and the receptor subtype. Cooperativity can be positive (allosteric modulator increases affinity for a given classical ligand), negative (allosteric modulator decreases affinity), or neutral (allosteric modulator binds to the receptor but does not change affinity for a given classical ligand). Mutagenesis studies have been used to reveal sensor sequences important for allosteric modulator affinity and cooperativity with classical ligands. These studies have used two different approaches, i.e., site-directed mutagenesis of specific receptor residues (Leppik et al., 1994; Matsui et al., 1995; Gnagey et al., 1999) or construction of hybrid receptors exploiting pharmacological differences among individual receptor subtypes (Ellis et al., 1993; Ellis and Seidenberg, 2000; Krejčí and Tuček, 2001). Together, experimental evidence is consistent with the notion that the second and the third extracellular loop mainly determine properties of interaction of allosteric ligands with muscarinic receptors.
In recent experiments, we exploited the divergence of allosteric interactions at M2 and M3 subtypes (Krejčí and Tuček, 2001). We found that complete exchange of the extracellular o2 loop (o2) of the M3 subtype of muscarinic receptor by the o2 loop of the M2 subtype increased affinity of the chimeric receptor (M3o2 chimera) for gallamine and alcuronium, but had no effect on their cooperativity with N-methylscopolamine (NMS) binding. Similar substitution of the extracellular o3 loop (M3o3 chimera) also increased affinity of the chimeric receptor for both allosteric modulators. Replacement of the o3 loop augmented negative cooperativity of gallamine and inverted negative cooperativity of alcuronium at parent M3 receptor to positive cooperativity on the donor M2 receptor. This was the first example of reversal of the direction of cooperativity that was accomplished by exchange of the defined segment of muscarinic receptor subtype displaying negative cooperativity for corresponding segment of a subtype exhibiting positive cooperativity (Krejčí and Tuček, 2001).
Using the same approach in the present experiments, we investigated the influence of similar and additional mutations in the third extracellular loop (o3) of the M3 receptor subtype on mutual interactions of the classical ligand NMS and four closely related derivatives of strychnine (Fig. 1). We demonstrate that the o3 loop of the M3o3 chimera is responsible for the appearance of positive cooperativity between NMS and Wieland-Gumlich aldehyde (WGA) or brucine. Results of our experiments establish for the first time that substitution of as little as three amino acids in the third extracellular loop of the M3 subtype of muscarinic receptor by corresponding residues of the M2 subtype brings about positive cooperativity between NMS and WGA, whereas this compound exerts negative cooperativity on both parent subtypes. We show in addition that positive cooperativity of tested strychnine-like modulators on the chimeric receptor is conditioned by proper interaction with the o2 loop.

Chemical structure of allosteric modulators.
Materials and Methods
Materials. WGA [(17R)-17,18-epoxy-17-hydroxy-19,20-didehydrocuranium] and propargyl-WGA [(4R,17R)-4-propargyl-17,18-epoxy-17-hydroxy-19,20-didehydrocuranium iodide] were synthesized by Dr. T. Elbert (Charles University, Prague, Czech Republic). [3H]NMS was from PerkinElmer Life and Analytical Sciences (Boston, MA). Brucine, strychnine, and atropine were from Sigma-Aldrich (Prague, Czech Republic).
Mutagenesis and Expression. Amino acids of the M3 receptor were always mutated to the corresponding residues of the M2 receptor (Fig. 2). Mutants of M3 receptor containing entire o2 or o3 loops of M2 sequence are named M3o2 and M3o3, respectively. For the sake of brevity, names of the rest of mutants consist of receptor subtype followed by a suffix of original amino acids in the third extracellular loop followed by the substituted amino acids. For example, M3SKFN→PNVT means that four amino acids (serine 519, lysine 523, phenylalanine 525, and asparagine 527) in the third extracellular loop of M3 receptor were mutated on proline, asparagine, valine, and threonine in M2 sequence (Fig. 2).

Sequences of amino acids in the second and third external loop of the M2 and M3 subtypes of human muscarinic receptors and nomenclature of the mutants. Data are taken from Bonner et al. (1988). Amino acids in the o2 or o3 extracellular loop of M3 receptor were mutated to the corresponding residue of M2 sequence. Chimeric M3 receptors incorporating entire sequence of o2 or o3 loop of M2 receptor are named M3o2 and M3o3, respectively. Accordingly, names of individual mutants consist of receptor subtype name followed by list of mutated amino acids in single letter code. For example, M3 K→N indicates that the lysine 523 of the third extracellular loop of the M3 receptor was mutated to the corresponding asparagine of M2 sequence, and M3 SKFN→PNVT means that four amino acids in the third extracellular loop of M3 receptor were mutated to their corresponding residues in M2 sequence.
Wild-type genes for human muscarinic M2 and M3 receptors incorporated in pCD vector (kindly provided by Professor N. Buckley, University Leeds, Leeds, UK) were used. Construction of M3 receptor with the substituted M2 o2 or o3 loop and mutations (M3K→N and M3KFN→NVT) in the o3 loop was described in Krejčí and Tuček (2001). Additional mutants were generated using QuikChange kit (Stratagene, La Jolla, CA). To replace serine 519 with proline, plasmid containing wild-type M3 receptor as template and 5′-TGA ACA CCT TTT GTG ACC CCT GCA TAC CCA AAA CCT TTT GG-3′ primer were used. To obtain M3DKFN→ANVT, mutant plasmid containing M3KFN→NVT mutant as template and 5′-GTG AAC ACC TTT TGT GCC AGC TGC ATA CCC AAT ACC-3′ primer were used. To obtain M3SK→PN, mutant plasmid containing M3 K→N mutant as template and 5′-TGA ACA CCT TTT GTG ACC CCT GCA TAC CCA ATA CCT TTT GG-3′ primer were used. To obtain M3SKFN→PNVT, mutant plasmid containing M3KFN→NVT mutant and 5′-TGA ACA CCT TTT GTG ACCCCT GCA TAC CCA ATA CCG TTT GG-3′ primer were used. All mutated DNAs were sequenced with the dideoxy method by Dr. J. Felsberg (Academy of Sciences, Institute of Microbiology, Prague, Czech Republic).
COS-7 cells were transiently transfected with the use of the DEAE-dextran method. Cells were grown in 10-cm Petri dishes in Iscove's modified Dulbecco's medium (Sigma-Aldrich) with 10% fetal calf serum. On day 1, 2 × 106 cells were seeded per dish. On day 3, after washing with phosphate-buffered saline, the transfection mix (2 μg of plasmid DNA and 0.27 mg of DEAE-dextran in 1 ml of phosphate-buffered saline) was applied for 2 h. Seven milliliters of serum-free Iscove's modified Dulbecco's medium and chloroquine in a final concentration of 80 μM were then added for an additional 3 h. Transfection medium was then removed, and fresh medium supplemented with 10% fetal calf serum was applied. Cells were harvested 72 h after transfection.
Radioligand Binding Experiments. Radioligand binding experiments were performed on membranes of COS-7 cells. Membranes were obtained by dilution of freshly harvested COS-7 cells in a medium composed of 136 mM NaCl, 5 mM KCl, 1 mM MgCl2, and 10 mM Na-HEPES, pH 7.4, to a final concentration of 107 cells per milliliter. Cell suspension was then homogenized (Ultra-Turrax; Janke & Kunkel GmbH & Co. KG, IKA-Labortechnik, Staufen, Germany) at 0°C by two 30-s strokes. Homogenate was centrifuged for 20 min at 30,000g. Pellets were resuspended in 10 times original volume (before centrifugation), and centrifugation was repeated. Membranes were kept frozen at –20°C for a maximum of 1 month and rehomogenized just before experiment. Binding experiments to membranes from 1 to 2 million cells per tube were done as described previously (Jakubík et al., 1995). The medium used for incubation was the same as described above, and incubation volume was 0.8 ml. Incubations were performed at 25°C and were terminated by filtration through Whatman GF/C glass fiber filters in a Brandel filtration apparatus (Sensat, Hatsfield, Hertshire, UK). Nonspecific binding was determined in the presence of 1 μM NMS.
The affinity of wild-type and mutated muscarinic receptors for [3H]NMS was measured in saturation binding experiments (1-h incubation at 25°C with [3H]NMS, at concentrations ranging from 25 pM to 1 nM) and expressed as Kd (equilibrium dissociation constant for the binding of [3H]NMS). The apparent dissociation constants, KA, of receptors for allosteric ligands and factors of cooperativity between allosteric ligand and [3H]NMS were determined in competition-type experiments in which membranes were preincubated in the presence of 200 pM [3H]NMS for 1 h at 25°C, and then increasing concentrations of the allosteric ligand were added and incubation was continued for an additional 5 h.
Dissociation constants koff in the absence and presence of allosteric ligands displaying neutral cooperativity with [3H]NMS were examined in a procedure modified from Lysíková et al. (1999). Membranes were preincubated for 1 h with [3H]NMS at a concentration corresponding to twice its Kd value. Brucine or WGA in concentrations ranging from 0.01 to 1 mM were added, and preincubation was continued for an additional 5 h. Dissociation was started by addition of NMS at a final concentration of 5 μM. Dissociation was stopped after 10 and 30 min, and koff values based on two-point determinations were computed for each concentration of the allosteric modulator.
Data Treatment. Data were processed and analyzed with open source software OpenOffice 1.0.2 (OpenOffice Foundation; www.openoffice.org) and Grace 5.1.12 (Grace Development Team; plasmagate.weizmann.ac.il/Grace) on Mandrake distribution of Linux (www.mandrake.com). The equations for nonlinear regression analysis were as follows.
Saturation binding experiments. After subtraction of nonspecific binding, eq. 1 was fitted to the data:  where Y is [3H]NMS binding at concentration X of free [3H]NMS, Kd is the equilibrium dissociation constant, and BMAX is the number of binding sites.
where Y is [3H]NMS binding at concentration X of free [3H]NMS, Kd is the equilibrium dissociation constant, and BMAX is the number of binding sites.
Competition-type experiments. After subtraction of nonspecific binding and normalization (to express the binding of [3H]NMS in the presence of allosteric ligand as percentage of the binding in the absence of allosteric ligand), eq. 2 was fitted to the data:  where Y is [3H]NMS binding at concentration of allosteric ligand X, N is concentration of [3H]NMS, Kd is the equilibrium dissociation constant obtained from saturation experiment, KA is apparent equilibrium dissociation constant of allosteric ligand, and α is the cooperativity factor between allosteric ligand and [3H]NMS (Ehlert, 1988).
where Y is [3H]NMS binding at concentration of allosteric ligand X, N is concentration of [3H]NMS, Kd is the equilibrium dissociation constant obtained from saturation experiment, KA is apparent equilibrium dissociation constant of allosteric ligand, and α is the cooperativity factor between allosteric ligand and [3H]NMS (Ehlert, 1988).
Dissociation experiments. After subtraction of nonspecific binding and normalization (to express the binding of [3H]NMS at a given time as percentage of the binding at time 0), eq. 3 was fitted to the data:  where Y is [3H]NMS binding at time X, and koff is rate dissociation constant.
where Y is [3H]NMS binding at time X, and koff is rate dissociation constant.
Results
Binding of [3H]NMS to Wild-Type Receptors and M3 Mutants. Plasmids containing cDNA of the M3 receptor were mutated in the region encoding the second (o2) or the third extracellular (o3) loops of the receptor so that the corresponding amino acid residue or residues (Fig. 2) of the M2 receptor were expressed. M2 wild-type (M2wt) and M3 wild-type (M3wt) receptors and all seven mutants of the M3 receptor were transiently expressed in COS-7 cells. Saturation binding experiments with [3H]NMS indicated that all constructs are expressed at a comparable level of 170 to 210 fmol of binding sites per milligram of protein (not shown). Although [3H]NMS is generally considered to be nonselective among muscarinic receptor subtypes, the affinity of M3 receptors for [3H]NMS (Kd = 0.32 ± 0.01; n = 6) expressed in COS-7 cells was approximately 2.4 times higher than affinity of M2 receptor (Kd = 0.78 ± 0.04; n = 6) (Table 1). This is in concert with data obtained using Chinese hamster ovary cells (Buckley et al., 1989; Lysíková et al., 1999) and COS-7 cells (Krejčí and Tuček, 2001). Mutation of three (M3KFN→NVT) or more residues in the o3 loop of the M3 receptor increased the Kd value for [3H]NMS, and exchange of the complete o3 loop led to a Kd value for [3H]NMS similar to that of the M2wt receptor. However, exchange of the complete second extracellular loop had no influence on affinity (Table 1). This is in agreement with a previous finding (Krejčí and Tuček, 2001).
Binding parameters of wild-type M2 and M3 receptors and modified M3 receptors
Equilibrium dissociation constants (Kd) for [3H]NMS were obtained from saturation experiments, and apparent equilibrium dissociation constants (KA) and factors of cooperativity α of allosteric modulators were obtained from competition-like arranged experiment with [3H]NMS as described under Materials and Methods. Data represent mean ± S.E.M of four to six or three independent experiments done in triplicates for wild-type and chimeric receptors and for receptors with mutations in the o3 loop, respectively.
Interaction of [3H]NMS and Allosteric Modulators. We tested four closely related chemicals: strychnine, its dimethoxy derivate brucine, strychnine synthesis precursor WGA, and its derivative propargyl-WGA as allosteric modulators of [3H]NMS binding to M2wt, M3wt, and mutated M3 subtypes of muscarinic receptors (Fig. 1). These compounds displayed a variety of cooperativities with [3H]NMS. Thus, there was relatively strong negative cooperativity on both wild subtypes by propargyl-WGA with weaker negative cooperativity by WGA on both subtypes. In contrast, brucine exhibited weak negative cooperativity on M3wt and positive cooperativity on M2wt receptors, whereas strychnine had positive cooperativity on both receptor subtypes (Table 1; Fig. 3). Tested modulators also differed in affinities for M2wt and M3wt receptors. Propargyl-WGA had 30 times higher affinity for the M2wt than for the M3wt receptor, whereas affinity of WGA was only 2 times higher at the M2 than the M3 receptor. Affinity of brucine was equal at both receptor subtypes, and the affinity of strychnine was slightly (2 times) lower on the M2 than on the M3 subtype (Table 1).

Modulation of [3H]NMS binding by strychnine (A), brucine (B), WGA (C), and propargyl-WGA (D) in M3wt, M2wt, and chimeric M3o2 and M3o3 receptors. Measurements of [3H]NMS binding and curve fitting were performed as described under Materials and Methods. Ordinate, [3H]NMS binding is expressed as percentage of control binding in the absence of indicated modulator, and each point represents mean ± S.E.M. of four to six independent experiments run in triplicates. Abscissa, concentration in log [M] of indicated allosteric modulator. Symbols denote M3 wild-type receptor (▪), M2 wild-type receptor (•), M3o3 chimeric receptor (▾), and M3o2 chimeric receptor (▴). Calculated binding parameters are presented in Table 1. A, replacement of the o2 extracellular loop in M3 receptor slightly increases binding of [3H]NMS in the presence of strychnine compared with M3wt, whereas replacement of the entire o3 extracellular loop results in virtually identical binding curve with M2wt receptor. B, replacement of the second extracellular loop of M3 receptor has no effect on interaction with brucine whereas M3o3 chimeric receptor exhibits strong positive cooperativity that is even about 3 times higher than at M2wt receptor. C, WGA exhibits weak negative cooperativity with [3H]NMS binding at both wild-type receptors. In contrast to M3o2 mutation, which has no effect on the interaction of WGA with [3H]NMS binding, M3o3 chimera displays significant positive cooperativity between WGA and [3H]NMS binding. D, propargyl-WGA interactions compared with M3wt display substantial increase of affinity and decrease of cooperativity at both M3o2 and M3o3 chimeric receptors that in the latter case even exceeds values of M2wt receptor.
Effects of Exchange of the Entire o2 or o3 Loop between M2 and M3 Receptors on Affinity and Cooperativity of Allosteric Modulators. Strychnine displayed positive cooperativity with [3H]NMS on both M2wt and M3wt receptors and had higher affinity for M3wt than for M2wt receptors. Exchange of the entire o2 loop of the M3wt receptor by the M2 sequence (M3o2 chimera) had no effect on the affinity of strychnine and slightly enhanced positive cooperativity between strychnine and [3H]NMS. Replacement of the entire o3 loop of the M3wt receptor with the M2 sequence (M3o3 chimera) also decreased affinity of strychnine below the value of M2wt receptor and enhanced positive cooperativity between strychnine and [3H]NMS. Resulting positive cooperativity on M3o3 chimera was significantly stronger than on M2wt receptor (Table 1; Fig. 3A).
Brucine had negative cooperativity with [3H]NMS binding on M3wt but positive cooperativity on M2wt. Both M3wt and M2wt receptors showed the same affinity for brucine. Exchange of the entire o2 loop had no effect either on affinity for brucine or cooperativity between brucine and [3H]NMS. Substitution of the entire o3 loop had no effect on affinity but reversed the negative cooperativity between brucine and [3H]NMS on parental M3wt receptor to positive cooperativity. As for strychnine, resulting positive cooperativity on chimeric M3o3 receptor was even stronger than on M2wt receptor (Table 1; Fig. 3B).
WGA exhibited negative cooperativity with [3H]NMS on both M3wt and M2wt receptors and had 2 times lower affinity for M3wt than for M2wt receptor. Exchange of the o2 loop had no effect on affinity of WGA or on its cooperativity with [3H]NMS. In contrast, exchange of the o3 loop increased affinity for WGA 4 times. Strikingly, the resulting M3o3 chimera exerted positive cooperativity between WGA and [3H]NMS, despite that parent M2wt as well as M3wt receptor demonstrated negative cooperativity. Because the negative cooperativity at the donor M2wt receptor is larger than that at the acceptor M3wt receptor, one would have expected enhanced negative cooperativity of WGA at the M3o3 chimera (Table 1; Fig. 3C).
Propargyl-WGA showed strong negative cooperativity on both M3wt and M2wt receptors and had more than 30 times lower affinity for M3wt than M2wt receptor. M3o2 chimera had more than 4 times higher affinity for propargyl-WGA than M3wt receptor. Cooperativity between propargyl-WGA and [3H]NMS on the M3o2 chimera was significantly weaker compared with both M3wt and M2wt receptors. Replacement of the o3 loop of M3 receptor brought about much stronger increase in affinity for propargyl-WGA than exchange of the o2 loop. Affinity of propargyl-WGA was about 40 times higher on M3o3 chimera than on M3wt and about 10 times higher than on M3o2 chimera. However, unlike other tested allosteric modulators, the effect of exchange of the o3 loop on cooperativity between propargyl-WGA and [3H]NMS was smaller than the effect of exchange of the o2 loop (Table 1; Fig. 3D).
Together, complete replacement of the M2o2 loop in the M3 receptor had no effect on the affinity of strychnine, brucine, and WGA. However, exchange of the o3 loop, with the exception of brucine, changed affinity of all other allosteric modulators. The general direction of these changes was always toward the values of the M2wt receptor, and in the case of strychnine and WGA, the affinity of the M3o3 chimera even surpassed that of the M2wt receptor. Exchange of the o3 loop affected cooperativity of binding with [3H]NMS of all tested allosteric modulators. In contrast, replacement of the o2 loop significantly influenced cooperativity with [3H]NMS only of strychnine and propargyl-WGA. Only propargyl-WGA showed stronger change of cooperativity at the M3o2 than the M3o3 chimera.
Effects of Specific Mutations in the o3 Loop. Because of the profound effects that exchange of the entire o3 loop of M3wt receptor has demonstrated on the binding characteristics of various allosteric modulators (Krejčí and Tuček, 2001; this report), the role of individual amino acids was studied in detail by their gradual mutations. We have started with two single mutations. Serine 519 was mutated to proline (M3S→P) and lysine 523 was mutated to asparagine (M3K→N). Subsequently, these mutations were combined (M3SK→PN), with or without additional mutations in the o3 loop of the M3 receptor (M3KFN→NVT, M3DKFN→ANVT, and M3SKFN→PNVT) to gradually match M2 sequence (for numbering of amino acids, see Fig. 2).
Strychnine had positive cooperativity with [3H]NMS on both subtypes and higher affinity for M3wt than M2wt receptor. As mentioned above, exchange of the entire o3 loop of M3wt receptor decreased affinity for strychnine below the value of the M2wt receptor and enhanced positive cooperativity with [3H]NMS above the value of M2wt receptor (Table 1; Figs. 3A and 4). Although none of the single mutations had comparable effect on affinity as replacement of the entire o3 loop, mutation of only one amino acid M3K→N (at position 523) significantly increased cooperativity of strychnine with [3H]NMS, and replacement of just two amino acids M3SK→PN (at positions 519 and 523) decreased affinity for strychnine in addition to increasing cooperativity to the value of M2wt receptor. Mutation of only three amino acids (lysine 523 to asparagine, phenylalanine 525 to valine, and asparagine 527 to threonine; M3KFN→NVT) augmented positive cooperativity between strychnine and [3H]NMS to the value of M2wt receptor, and additional mutations to this triple-mutant further increased this positive cooperativity.

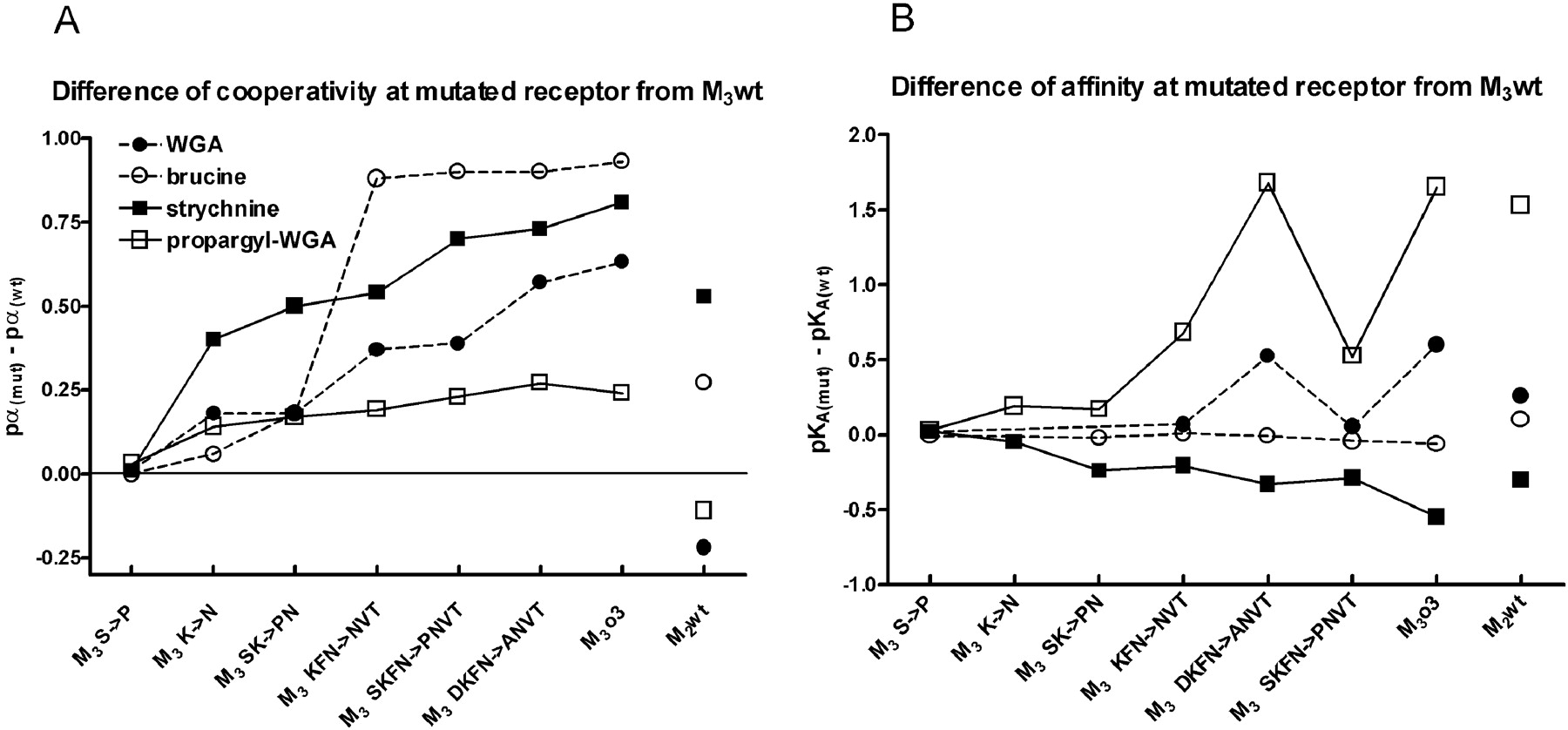
Differences of allosteric modulator affinity and cooperativity with [3H]NMS binding between M3 wt and mutated receptors. Divergence of binding parameters was calculated by subtracting the experimentally determined values listed in Table 1 at mutated receptors from those of host M3 wt receptor. Statistical significance and number of observations are as indicated in Table 1. Symbols denote strychnine (▪), propargyl-WGA (□), brucine (○), and WGA (•). Abscissa, gradational mutations in the o3 extracellular loop of M3wt receptor are labeled as described under Materials and Methods (Fig. 2; Table l). A, ordinate, differences of cooperativity between [3H]NMS and allosteric modulators are given as differences of pα values. Note that all changes of cooperativities are positive. B, ordinate, changes of affinities of allosteric modulators are given as differences of pKA values. Note that there is no clear correlation between changes of cooperativity and affinity. For comparison, differences M2wt versus M3wt are plotted on the right-hand side (M2wt).
Brucine displayed negative cooperativity on M3wt receptor and positive cooperativity on M2wt receptor and has the same affinity for both subtypes. Exchange of the entire o3 loop of M3wt receptor with M2 sequence did not change affinity for brucine, but it reversed negative cooperativity between brucine and [3H]NMS to positive cooperativity that was even higher than on the donor M2wt receptor (Table 1; Figs. 3B and 4). Mutation of just two amino acids (M3SK→PN) was sufficient to change negative cooperativity to positive cooperativity, and mutation of three amino acids (M3KFN→NVT) had the same effect as exchange of the entire o3 loop. Due to the neutral cooperativity between brucine and [3H]NMS, affinity of M3K→N mutant for brucine was determined in dissociation experiments. Mutant M3K→N has similar affinity to M3wt (pKA = 3.99 and 4.00 in two independent experiments performed in triplicates; data not shown).
WGA exhibited negative cooperativity on both M2wt and M3wt receptor and had 2 times higher affinity for M2wt than for M3wt receptor. Exchange of the entire o3 loop of M3 receptor with M2 sequence increased affinity 4 times to a value significantly higher than that of M2wt receptor and changed cooperativity between WGA and [3H]NMS from negative to positive (Table 1; Figs. 3C and 4). Single mutation of M3K→N changed negative cooperativity to neutral and mutation of three amino acids M3KFN→NVT changed negative cooperativity of M3wt receptor to positive without changing affinity for [3H]NMS. Replacement of four amino acids M3DKFN→ANVT brought about an increase of affinity for WGA that became higher than at M2wt receptor and further enhanced positive cooperativity so that both affinity and cooperativity of WGA equaled that of M3o3 chimera. Due to neutral cooperativity between WGA and [3H]NMS, affinity of M3K→N and M3SK→PN mutants for WGA was determined in dissociation experiments. Mutant M3K→N (pKA = 3.93, 3.95) as well as M3SK→PN (pKA = 3.99, 4.00; two independent experiments in triplicates; data not shown) had lower affinity than M3wt. However, it should be noted that affinities of allosteric ligands determined by dissociation experiments do not always strictly correspond to those determined in competition-type experiments (Krejčí and Tuček, 2001).
Similar to WGA, propargyl-WGA displayed negative cooperativity with [3H]NMS on both M3wt and M2wt receptor that is stronger on M3wt receptor. M2wt receptor had 30 times higher affinity for propargyl-WGA than M3wt receptor (Table 1; Figs. 3D and 4). Exchange of the entire o3 loop of M3 receptor increased affinity for propargyl-WGA and reduced negative cooperativity between propargyl-WGA and [3H]NMS. Similar to WGA, the orientation of change in cooperativity was in the opposite direction than at wild subtypes, but, unlike WGA, it did not reach positive values. Mutation of just one amino acid (M3K→N) made cooperativity between propargyl-WGA and [3H]NMS significantly weaker and increased affinity for propargyl-WGA (Table 1; Fig. 4, A and B). Replacement of four amino acids M3DKFN→ANVT resulted in change of affinity for propargyl-WGA as well as of its cooperativity with [3H]NMS to the value at M3o3 chimera.
Discussion
We took advantage of the known stronger allosteric interactions on M2 than on M3 receptor subtype to investigate a potential role of cognate amino acid residues in the third extracellular loop that differ between the two subtypes in positive cooperativity of some strychnine-like allosteric modulators with [3H]NMS. The most important finding of our study is discovery of positive cooperativity between WGA and [3H]NMS on chimeric M3o3 receptor constructed by substitution of the o3 loop of M3wt receptor by that of M2wt receptor. This is in spite the fact that WGA has negative cooperativity with [3H]NMS on both wild receptor subtypes. To our knowledge, this is the first evidence of manifestation of reversal of the direction of a negative cooperativity that is based on exchange of amino acid sequences involved in its binding. This result clearly demonstrates that modifications in the o3 loop are propagated to other parts of the receptor where new intramolecular interactions may occur. Spreading of the conformational change is further supported by the decrease of affinity for the orthosteric antagonist [3H]NMS (Table 1) whose binding site is located deep within transmembrane segments. This notion is also consistent with the finding of Lee at al. (1992) who showed that mutation of aspartate 71 in the second transmembrane segment of the M1 receptor, an amino acid far away from the allosteric binding site, decreases affinity for the allosteric modulator gallamine. Similarly, changes of NMS dissociation rate from M2 receptor modified in the second and the third extracellular loops has been reported (Buller et al., 2002; Voigtländer et al., 2003). Although muscarinic receptors are polytopic membrane proteins consisting of independent domains, the o3 loop cannot be considered an independent domain (Popot and Engelman, 1990; Maggio et al., 1993a,b; Novi et al., 2003). Therefore, observed effects of mutations in the o3 loop on cooperativity and affinity of the allosteric binding site should not be interpreted as indicative of a sole role of the o3 loop.
Our data point to an essential role of the o3 loop in the observed reversal of negative cooperativity between NMS and WGA or brucine. Replacement of the o2 loop in the M3wt receptor does not influence either affinity for WGA or cooperativity of WGA with NMS, or affinity for NMS. In the case of brucine that has the same affinity for both wild receptor subtypes, complete replacement of the o2 or the o3 extracellular loops and any mutations in the o3 loop expectedly do not influence its affinity. Similar to WGA, the exchange of the o2 loop has no influence on moderate negative cooperativity of brucine with NMS at M3wt receptor. Although tested strychnine-like allosteric modulators display both negative and positive cooperativity at M2wt receptor, the transposition of its o3 loop to M3wt receptor in all cases shifts cooperativity in one direction (Fig. 4A), i.e., decreases original negative or increases positive cooperativity. Out of five amino acids that differ between the o3 loops of the two receptors, the most important amino acids of the o3 loop of M2 receptor for this shift to become apparent are asparagine, valine, and threonine. Participation of these residues that are close to the seventh transmembrane domain corroborate the importance of this segment in allosteric interactions, namely, in determining affinity of allosteric modulator binding (Matsui et al., 1995; Buller et al., 2002; Voigtländer et al., 2003). However, in this context it is worth noting that the pattern of changes of cooperativity between [3H]NMS and strychnine-like allosteric modulators induced by mutations does not overlap with the pattern of their changes of affinity, e.g., gradual decrease of affinity of strychnine is accompanied by increase of cooperativity. Similarly, large increase of cooperativity of brucine is not associated with changes of affinity, and the large decrease of affinity for propargyl-WGA observed between mutants M3DKFN and M3SKFN is not accompanied with change of cooperativity (Fig. 4). This lack of correlation further demonstrates that amino acid residues essential for transmission of conformational change are not necessarily the same as those that form the binding site of allosteric modulators. A common transduction mechanism of cooperativity changes of all tested strychnine-like modulators is further supported by similar pattern of effects of gradual mutations (Fig. 4A)
The divergence of allosteric properties between strychnine, brucine, and WGA on one side and propargyl-WGA on the other side may be due to a difference in the locations to which they bind. Exchange of the entire o2 loop of the M3wt receptor with M2 sequence has no influence on affinity for NMS and thus in itself, unlike replacement of the o3 loop, does not induce a conformational change propagated to the orthosteric binding site. This modification, however, increases affinity for propargyl-WGA, hence indicating difference in binding of this allosteric modulator with the o2 loop of M2 and M3 receptors. The important role of the o2 loop of M2 receptor in the affinity for alcuronium, a compound structurally similar to a dimer of propargyl-WGA, and gallamine, a compound structurally dissimilar to tested strychnine-like modulators, was demonstrated (Gnagey et al., 1999; Krejčí and Tuček, 2001). Similarly, interaction of these ligands with the o2 and the o3 loop of muscarinic M3 receptor was shown (Jakubík and Wess, 1999). Involvement of the o2 loop in high-affinity binding of propargyl-WGA on M2 receptor is in line with reported role of the o2 loop together with tyrosine 423 in the o3 loop in high-affinity binding of structurally different caracurine derivatives and alkane-bisammonium compounds to M2 receptor (Buller et al., 2002; Voigtländer et al., 2003). Together, these findings are compatible with and further support the view of muscarinic receptors as proteins with multiple allosteric binding sites (Lazareno et al., 2000; Birdsall et al., 2001).
Results of our experiments seem to be explained most easily on the assumption that an interplay of the o2 and the o3 extracellular loops plays a specific role in a conformational shift induced by binding of tested strychnine-like allosteric modulators and resulting cooperativity with [3H]NMS. WGA and brucine, whose binding to the o2 loop does not differ between M2wt and M3wt, display positive cooperativity with NMS at chimeric M3o3 receptor regardless of cooperativity they have on parent wild-type receptors. Strychnine and propargyl-WGA, whose binding is affected by exchange of the o2 loop, show more positive cooperativity at M3o3 in comparison with M3wt when parent receptors have positive cooperativity (strychnine) or less negative cooperativity when parent receptors have negative cooperativity (propargyl-WGA). Replacement of the o2 loop (chimera M3o2) slightly but significantly increases positive cooperativity of strychnine that is, however, much weaker than its positive cooperativity at M3o3 chimera. These observations suggest that the interaction of allosteric ligands with the o3 loop that was transposed from M2wt to create M3o3 chimera always induces conformational change, manifesting itself as positive cooperativity provided that there is no difference in interaction of allosteric modulator with o2 loop of M3 and M2 receptors as in the case of strychnine, brucine, and WGA. Different binding of propargyl-WGA (exhibiting strong negative cooperativity at parent wild-type receptors) to o2 loop of M3 and M2 receptor impedes this presumed conformational change and results in only reduction of negative cooperativity at M3o3 chimera. In line with this notion are results of experiments using M3o2 chimeric receptors. Complete replacement of the o2 loop of the M3wt receptor with that of M2wt does not change affinity for strychnine, brucine, and WGA and has little or no effect on cooperativity with NMS. On the other hand, propargyl-WGA displays higher affinity and a decrease in negative cooperativity with NMS at M3o2 chimera, which is significantly more pronounced than that at M3o3 chimera (more than 2 times).
In conclusion, the positive cooperativity of studied strychnine-like allosteric modulators with the binding of [3H]NMS is mainly determined by the o3 loop of the receptor, namely, by three amino acids—asparagine, valine, and threonine—located close to the seventh transmembrane domain of M2wt receptor. Their incorporation into the o3 loop of M3 receptor leads to manifestation of positive cooperativity between WGA and [3H]NMS, despite that both M2wt and M3wt receptors exert negative cooperativity. The reversal of original negative cooperativity at M3wt receptor to positive at M3o3 chimera is confined to WGA and brucine. Propargyl-WGA that has similarly to chemically closely related WGA negative cooperativity at both wild-type receptors does not exhibit this switch. Our data suggest that different epitopes on the outward oriented surface of muscarinic receptor are involved in binding of different allosteric modulators, whereas structures underlying the mechanism of allosteric interaction between allosteric modulators and orthosteric antagonist are similar. The dissimilarity of WGA and propargyl-WGA in binding to receptor is most likely due to the propargyl group and quaternary nitrogen (Fig. 1) that are apparently involved in interaction with the o2 loop and lead to high affinity for M2wt. However, differences in chemical structure among other tested strychnine-like modulators responsible for distinct allosteric interactions are not clear. To clarify this issue, more studies of allosteric modulators of various chemical structures and fitting structures of these modulators to a molecular model of muscarinic receptor will be required.
Acknowledgments
We appreciate the vital contribution of deceased Prof. H. C. Stanislav Tuček in elaboration of the project and the most valuable help of Profs. E. E. El-Fakahany and N. J. Buckley.
Footnotes
-
This work was supported by research project AVOZ 50110509, Grant Agency of the Czech Republic Grants 309/02/1331 (to V.D.) and 305/02/D090 (to J.J.), and Grant Agency of the Academy of Sciences of the Czech Republic Grant A5011306 (to V.D.).
-
doi:10.1124/jpet.104.080358.
-
ABBREVIATIONS: NMS, N-methylscopolamine; WGA, Wieland-Gumlich aldehyde; o2, second extracellular loop of receptor; o3, third extracellular loop of receptor; M3o2, chimeric receptor composed of M3 receptor with the o2 loop of M2 receptor; M3o3, chimeric receptor composed of M3 receptor with the o3 loop of M2 receptor; wt, wild-type.
- Received November 8, 2004.
- Accepted January 11, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}