


Abstract
Sch527123 [2-hydroxy-N,N-dimethyl-3-[[2-[[1(R)-(5-methyl-2-furanyl)propyl]amino]-3,4-dioxo-1-cyclobuten-1-yl]amino]ben-zamide] is a potent, selective antagonist of the human CXCR1 and CXCR2 receptors (Gonsiorek et al., 2007). Here we describe its pharmacologic properties at rodent CXCR2 and at the CXCR1 and CXCR2 receptors in the cynomolgus monkey, as well as its in vivo activity in models demonstrating prominent pulmonary neutrophilia, goblet cell hyperplasia, and mucus production. Sch527123 bound with high affinity to the CXCR2 receptors of mouse (Kd = 0.20 nM), rat (Kd = 0.20 nM), and cynomolgus monkey (Kd = 0.08 nM) and was a potent antagonist of CXCR2-mediated chemotaxis (IC50 ∼3–6 nM). In contrast, Sch527123 bound to cynomolgus CXCR1 with lesser affinity (Kd = 41 nM) and weakly inhibited cynomolgus CXCR1-mediated chemotaxis (IC50 ∼1000 nM). Oral treatment with Sch527123 blocked pulmonary neutrophilia (ED50 = 1.2 mg/kg) and goblet cell hyperplasia (32–38% inhibition at 1–3 mg/kg) in mice following the intranasal lipopolysaccharide (LPS) administration. In rats, Sch527123 suppressed the pulmonary neutrophilia (ED50 = 1.8 mg/kg) and increase in bronchoalveolar lavage (BAL) mucin content (ED50 =<0.1 mg/kg) induced by intratracheal (i.t.) LPS. Sch527123 also suppressed the pulmonary neutrophilia (ED50 = 1.3 mg/kg), goblet cell hyperplasia (ED50 = 0.7 mg/kg), and increase in BAL mucin content (ED50 = <1 mg/kg) in rats after i.t. administration of vanadium pentoxide. In cynomolgus monkeys, Sch527123 reduced the pulmonary neutrophilia induced by repeat bronchoscopy and lavage (ED50 = 0.3 mg/kg). Therefore, Sch527123 may offer benefit for the treatment of inflammatory lung disorders in which pulmonary neutrophilia and mucus hypersecretion are important components of the underlying disease pathology.
Chronic obstructive pulmonary disease (COPD) is the fourth major cause of death in the United States and is characterized by irreversible airflow limitation due to chronic bronchitis or emphysema (Barnes and Stockley, 2005). The pathological hallmarks of COPD include peripheral airway inflammation dominated by neutrophils, the destruction of the lung parenchyma, submucosal gland hypertrophy, and goblet cell hyperplasia, as well as an increase in proinflammatory cytokines and chemokines (Barnes and Stockley, 2005). Current therapies for COPD are similar to those for asthma and include the use of β-adrenoceptor agonists, theophylline, muscarinic antagonists, and corticosteroids (Barnes and Stockley, 2005). However, these treatments do not prevent the progressive decline in lung function and demonstrate clinical activity in only a subpopulation of COPD patients. There is a strong correlation between disease severity, exacerbations, and neutrophil infiltration into the pulmonary tissues of COPD patients (Stockley, 2002), and gene profiling of peripheral blood neutrophils from COPD subjects has indicated a statistically significant increase in the expression of several inflammatory genes compared with controls (Oudijk et al., 2005). Therefore, therapies directed at controlling pulmonary neutrophilia have been suggested as having potential benefit in COPD patients (Barnes and Stockley, 2005). One mechanism by which exacerbated neutrophil recruitment may mediate this disease is by the production of elastases, such as matrix metalloprotease-9 (MMP-9) (Kim and Nadel, 2004; Barnes and Stockley, 2005), although some investigators have reported an inverse correlation between neutrophil elastase levels and airspace changes (Eidelman et al., 1990).
The CXC glutamine-leucine-arginine motif (ELR) chemokines may be central to the inflammatory response in COPD. Both the CXCR1 and CXCR2 receptors are expressed in high numbers on human neutrophils (Mukaida, 2002) and have been detected on other cells associated with chronic inflammation, including macrophages, lymphocytes, mast cells, dendritic cells, and endothelium (Mukaida, 2002; Traves et al., 2004; Reutershan et al., 2006). Several ELR chemokines serve as high-affinity ligands for these receptors, with interleukin-8 (CXCL8) selectively acting through the human and cynomolgus monkey CXCR1 receptor and the CXCR2 receptor responding to other ELR chemokines, such as growth-related protein-α (GRO-α; CXCL1), neutrophil-activating peptide-2 (CXCL7), and epithelial-derived neutrophil attractant-78 (CXCL5), as well as CXCL8 (Jones et al., 1996; Mukaida, 2002; Hipkin et al., 2004). This chemokine network has been suggested to be important to the development of the neutrophilic lung inflammation, which exists in COPD (Barnes and Stockley, 2005), asthma, adult respiratory distress syndrome (Pease and Sabroe, 2002), and cystic fibrosis (Davis et al., 1996). Indeed, the levels of CXCL8 but not CXCL5 in the bronchoalveolar lavage fluid of cigarette smokers with emphysema were significantly elevated (Tanino et al., 2002), whereas a significant up-regulation of CXCL5, CXCL8, CXCR1, and CXCR2 gene expression was reported in endobronchial biopsies taken from COPD patients with exacerbations relative to patients with stable disease or non-smokers (Qiu et al., 2003). However, the lack of marked efficacy of a humanized anti-CXCL8 antibody in COPD patients (Mahler et al., 2004) indicates that the selective targeting of CXCL8 alone is not sufficient for clinical activity in this patient population and suggests that a therapeutic approach that targets the activity of multiple ELR ligands may be more suitable. One such approach would involve the selective antagonism of the CXCR1 and/or CXCR2 receptors. Previously, the activity of a CXCR2 antagonist Sch-N in a murine model of pulmonary neutrophilia induced by cigarette smoke inhalation has been described previously (Thatcher et al., 2005). The nomenclature of Sch-N has now been modified to Sch527123, and we now have extensively evaluated its activity in a series of murine, rat, and cynomolgus monkey models of pulmonary inflammation that recapitulate many of the critical features of COPD. Our results indicate that Sch527123 represents a novel approach for the treatment of pulmonary disorders characterized by pro-nounced pulmonary neutrophil trafficking, mucus hypersecretion, and goblet cell hyperplasia.
Materials and Methods
Materials. Sch527123 [2-hydroxy-N,N-dimethyl-3-{2-[[(R)-1-(5methyl-furan-2-yl)-propyl]amino]-3,4-dioxo-cyclobut-1-enylamino}-benzamide] was synthesized at the Schering-Plough Research Institute (Kenilworth, NJ). Lipopolysaccharide (LPS) from Escherichia coli was purchased from Sigma Chemical Co. (St Louis, MO), and vanadium pentoxide (V2O5) was purchased from Aldrich Chemicals (Milwaukee, WI).
Radioligand Binding Assays. [3H]Sch527123 was synthesized as described previously (Gonsiorek et al., 2007). Saturation bindings assays were done using scintillation proximity assay technology as described previously (Gonsiorek et al., 2007), using membranes prepared from murine BaF/3 cells stably transfected with murine, rat, or cynomolgus monkey CXCR1 or CXCR2.
Chemotaxis Assays. These assays were performed as described previously (Hipkin et al., 2004; Gonsiorek et al., 2007), using murine BaF/3 cells stably transfected with murine, rat, or cynomolgus monkey CXCR1 or CXCR2. Recombinant mouse macrophage inhibitory protein-2 (MIP-2) was used to induce mouse CXCR2-mediated chemotaxis; recombinant rat cytokine-induced neutrophil chemoattractant-3 (CINC-3) was used to induce rat CXCR2-mediated chemotaxis, and human recombinant interleukin-8 was used to stimulate cynomolgus monkey CXCR1- and CXCR2-mediated responses. All chemokines were purchased from R&D Systems (Minneapolis, MN). The reported IC50 values for Sch527123 were determined in the presence of 3 nM of the relevant chemokine.
Animal Care and Use. These experiments were performed with the prior approval of the Animal Care and Use Committee of Schering-Plough Research Institute, which is a facility accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International.
LPS Challenge in Mice. Male BALB/c mice (Jackson Laboratories, Bar Harbor, ME) weighing between 20 and 25 g were anesthetized by intraperitoneal injection of ketamine (100 mg/kg) and xylazine (10 mg/kg), and 50 μl of a 1 mg/ml LPS solution (50 μg/mouse) was instilled into the lungs via the intranasal route of administration. Control mice received intranasal injection of 50 μl of isotonic (0.9%) saline. The animals were allowed to recover under a heat lamp and returned to their cages. The next day, the procedure was repeated to administer a second dose of intranasal LPS or saline. Twenty-four hours after the second LPS administration, the mice were euthanized, a tracheal catheter was inserted, and a bronchoalveolar lavage (BAL) sample and lung tissues were collected. For the BAL collection, two 0.3-ml aliquots of isotonic saline were injected into the tracheal catheter, and the recovered fluid (approximately 80% recovery) was analyzed for total and differential cell analysis (Kung et al., 1994). Total cell counts were performed by hemocytometry. Slides for differential cell enumeration were prepared on a Shandon cytospin (Shandon, Pittsburgh, PA) at 250 rpm for 10 min, fixed with methanol, and stained with Hema 3 (Fisher Scientific, Pittsburgh, PA). At least 200 cells were counted from each cytospin to generate data. A separate set of animals was used to study the efficacy of Sch527123 on LPS-induced histopathological changes in the lungs. In this study, the highest dose of Sch527123 tested was 3 mg/kg. For the histological evaluation, the lungs were perfused in situ by flushing the pulmonary vasculature with buffered saline and fixed in 10% phosphate-buffered formalin. The right lobe of the lung was mounted in paraffin, and 5-μm sections were cut and stained with both hematoxylin and eosin (H&E), as well as periodic acid Schiff's (PAS) stains. The quantification of goblet cells was performed with an image analysis system that measures the percentage of PAS-staining positive cells within the airway epithelium (Kung et al., 1994). This analysis was performed on five randomly selected airways. In addition, H&E sections were examined for a qualitative assessment of pulmonary neutrophilic inflammation in the vicinity of airway epithelium and the alveoli.
Sch527123 was suspended in 0.4% methylcellulose and given orally by gavage 2 h before and 4 h after each intranasal administration of LPS. Control animals received 0.4% methylcellulose (10 ml/kg). In total, four doses of Sch527123 or vehicle were given.
LPS Challenge in Rats. Male Sprague-Dawley rats (200 g) from Charles River Laboratories (Wilmington, MA) were anesthetized with 5% isoflurane supplemented with oxygen and received 100 μlof LPS (100 μg/ml), dissolved in isotonic (0.9%) saline to deliver a dose of 10 μg/rat. Control animals received 100 μl of isotonic saline. The LPS or saline was delivered directly to the trachea with the aid of a fine-tipped microsprayer (Penn-Century, Philadelphia, PA) and laryngoscope. After recovery from the intratracheal injections, the rats were returned to their cages and given free access to food and water. Twenty-four hours after the LPS or vehicle challenge, the rats were euthanized, a tracheal catheter was inserted, and a BAL fluid sample was obtained. The BAL sample was analyzed for total and differential inflammatory cell numbers, for measurement of mucin content using the monoclonal anti-mucin antibody A10G5 (Phillips et al., 2006), and for the measurement of CINC-2α and CINC-3 by ELISA (IBL-America, Minneapolis, MN). The details for the LPS challenge and BAL cell enumeration have been described previously (Spond et al., 2001).
Sch527123 was suspended in 0.4% methylcellulose vehicle and given orally 2 h before the LPS challenge. Control rats received oral methylcellulose (10 ml/kg). Only one dose of Sch527123 or vehicle was given in these experiments.
V2O5 Challenge in Rats. Male Sprague-Dawley rats (Charles River Laboratories) weighing approximately 200 g were prepared for intratracheal injections as described above for the LPS challenge. Vanadium pentoxide (1 mg/ml) was dissolved in isotonic (0.9%) saline, and each rat received either 200 μl of V2O5 (1 mg/kg) or saline. For studies involving the assessment of pulmonary neutrophilia after V2O5 challenge, Sch527123 or 0.4% methylcellulose vehicle was given for 3 consecutive days starting 1 day and 1 h before the V2O5 challenge, followed by a third dose 1 day after the V2O5 challenge. The rats were euthanized on the following day, i.e., 48 h after the V2O5 challenge, and the total and differential cell counts in the BAL fluid were measured as described previously (Spond et al., 2001). A similar dosing paradigm was used for the studies involving goblet cell measurement only; Sch527123 or vehicle was given for eight consecutive daily doses. Sch527123 or vehicle was given 1 day and 1 h before the V2O5 challenge followed by daily administration of one dose per day for the next 6 days. The rats were euthanized on the following day, 7 days after the V2O5 challenge, and goblet cell numbers per unit area of airway epithelium was determined by image analysis as described above for studies involving LPS challenge in mice. The percentage of PAS+ cells and mucin content in the BAL fluid were determined as described above.
Repeat Bronchoscopy and Lung Lavage in Primates. Twenty-four male cynomolgus monkeys were used for these studies. In one experiment, six monkeys were orally dosed with Sch527123 (3 mg/kg), and six monkeys were dosed with 0.4% methylcellulose vehicle. In another experiment, six monkeys were orally dosed with Sch527123 (0.3 mg/kg), and the remaining six monkeys received the methylcellulose vehicle.
Each animal was placed in a restraining chair and dosed by oral gavage with Sch527123 or the vehicle administered in a volume of 4 ml followed by a 4-ml flush with methylcellulose and 3 ml of air. The oral dosing was performed with a stomach tube. The monkeys were left in the restraining chair and 2 h later were sedated with i.m. ketamine. A catheter was introduced into the cephalic vein, and anesthesia was induced by intravenous propofol (1–2 ml bolus; 2–4 mg/kg). Blood pressure, heart rate, and arterial oxygen saturation were monitored using techniques that have been described previously (Young et al., 1999). Before the introduction of the bronchoscope, a 2- to 3-ml bolus of propofol was given to suppress the cough reflex. The monkey was then placed in the supine position, and the mouth was opened with a laryngoscope. A small diameter (i.d. = 1.7 mm) polyethylene catheter was then inserted into the trachea through which a steady flow of 100% O2 was produced. An Olympus BF160 bronchoscope (Olympus Inc., Tokyo, Japan) was then inserted into the trachea and right main stem bronchus and wedged into a lobar bronchus. Images of the airway were recorded with a computerized videoprocessor. With the bronchoscope wedged into the airway, a BAL was attained by instilling and then withdrawing 2 × 10-ml aliquots of 0.9% saline. The BAL fluid was kept on ice until processed. The bronchoscope was then withdrawn from the airway, the propofol infusion was discontinued, and the monkeys recovered and then were returned to their cages.
Twenty-four hours later, the monkeys were anesthetized, and the bronchoscope was inserted into the same lobar bronchus used for the BAL collection at the 2-h time point. The location of the bronchoscope was verified using the airway images previously obtained with the videoprocessor. At this point, a BAL fluid sample was obtained. The BAL samples were analyzed for the total and differential cell enumeration as described previously (Young et al., 1999). Interleukin-8 levels were measured in the BAL fluid by ELISA following the manufacturer's protocol (R&D Systems, Inc., Minneapolis, MN).
Statistical Analysis. Data are presented as mean ± S.E.M., with statistical significance (p < 0.05) determined between groups using an analysis of variance and a post hoc analysis of Fisher's protected least square difference on a program from Stat-View (version 4.0; ABACUS Concept, Berkeley, CA).
Results
Inhibition of CXCR1 and CXCR2-Mediated Chemotaxis by Sch527123 in Rodents and Monkeys. Saturation binding analysis with [3H]Sch527123 was initiated as described previously (Gonsiorek et al., 2007) using membranes from BaF/3 cells stably transfected to express mouse CXCR2, rat CXCR2, or cynomolgus monkey CXCR1 and CXCR2. Membranes were incubated with increasing concentrations of [3H]Sch527123 in the presence or absence of excess unlabeled compound. As seen in Table 1, Sch527123 binds with high-affinity binding to CXCR2 in all species, especially cynomolgus CXCR2. The affinity for cynomolgus CXCR1 is lower than what was measured for human CXCR1 (Gonsiorek et al., 2007). Cells expressing these receptors were induced to migrate across a chemokine gradient in the presence or absence of a concentration range of Sch527123. As indicated in Table 2, Sch527123 inhibits CXCR2-mediated chemotactic responses from each of these species with an IC50 of ∼3 to 6 nM, similar to that observed versus human CXCR2 (Gonsiorek et al., 2007). In accordance with the saturation binding assays, Sch527123 is markedly less potent at the cynomolgus CXCR1 versus the human homolog (Table 1 compared to data presented in Gonsiorek et al., 2007).
Binding affinities of Sch527123 at mouse, rat, and monkey CXCR2 and CXCR1
All values expressed as mean nanomolar ± S.E.M.
Inhibition of CXCR1- and CXCR2-mediated chemotaxis by Sch527123
Sch527123 Is Orally Bioavailable in Mice, Rats, and Monkeys. Oral administration of Sch527123 results in substantial plasma levels in mice, rats, and cynomolgus monkeys (Table 3). These data indicate that Sch527123 concentrations in the plasma of animals are adequate for in vivo pharmacology studies in these species.
Pharmacokinetic parameters following a single oral dose of Sch527123
LPS Challenge in Mice. Treatment of mice with Sch527123 (0.1–10 mg/kg p.o.), given 2 h before and 4 h after each LPS challenge, inhibited the influx of neutrophils and total cells in the BAL fluid in a dose-dependent manner (Fig. 1). The ED50 of Sch527123 for inhibition of the pulmonary neutrophil influx induced by LPS challenge in these mice was 1.2 mg/kg. Sch527123 (0.1–3 mg/kg) also attenuated the increase in PAS staining in the airway epithelium following LPS challenge (Figs. 1 and 2). Statistically significant effects were seen at oral doses of Sch527123 of 1 and 3 mg/kg that produced a 32 ± 6 and 38 ± 6% inhibition of the PAS staining, respectively (Fig. 1). Histological evaluation of the lung tissue confirmed that LPS challenge produced goblet cell hyperplasia in the airway epithelium (Fig. 2, A and B), which was partially inhibited by Sch527123 (3 mg/kg p.o) (Fig. 2C). LPS challenge also produced a marked increased in the alveolar cellularity consisting predominantly of neutrophils (Fig. 2, D and E), which was partially inhibited by Sch527123 (3 mg/kg p.o.) (Fig. 2, F and G).
LPS Challenge in Rats. Sch527123 (0.1–3 mg/kg p.o.) dose-dependently reduced the number of neutrophils and total cells appearing in the BAL fluid 24 h after the LPS challenge (Fig. 3). The ED50 of Sch527123 for inhibition of the LPS-induced pulmonary neutrophilia in rats was 1.8 mg/kg. Treatment of rats with Sch527123 (0.1–1 mg/kg) also reduced the increase in BAL mucin content induced by the LPS challenge (Fig. 3). The ED50 for inhibition of the increased BAL mucin content was <0.1 mg/kg with all three doses producing near complete inhibition.
LPS challenge increased the levels of CINC-2α and CINC-3 in the BAL fluid. These chemokines seemed to peak at 4 h after LPS challenge and then returned close to baseline levels by 24 h (Fig. 4). In the presence of Sch527123 (3 mg/kg p.o.), CINC levels remained elevated throughout 24 h, at which point they were significantly increased relative to values in animals that received vehicle (Fig. 4).
V2O5 Challenge in Rats. Intratracheal challenge of rats with V2O5 increased the number of neutrophils and mucin content in the BAL fluid and the increased the percentage of PAS staining in the airway epithelium (Fig. 5). Treatment of rats with Sch527123 (1 and 3 mg/kg p.o.) reduced the number of BAL neutrophils and mucin content and attenuated the increase in PAS staining induced by the V2O5 challenge (Fig. 5). A second study was performed to expand the dose-response data with Sch527123, and the combined results from these two studies produced an ED50 of 1.3 mg/kg for the inhibition of the pulmonary neutrophilia and an ED50 of 0.7 mg/kg for the inhibition of the PAS staining. The ED50 of Sch527123 for inhibition of the increase in BAL mucin was <1 mg/kg.

Attenuation of neutrophil recruitment and goblet cell hyperplasia by Sch527123 treatment following intranasal LPS challenge of mice. A, total cells per milliliter of bronchoalveolar lavage fluid (BALF). B, number of neutrophils per milliliter of BALF. C, percentage of PAS-staining positive cells within the airway epithelium. Data represent the mean ± S.E.M. of 5 to 15 animals per treatment group. *, p < 0.05 compared to LPS + vehicle group.
Repeat Bronchoscopy and Lung Lavage in Primates. Repeat bronchoscopy and lung lavage, performed twice over a 24-h period led to an increase in the number of total cells and neutrophils recovered from the BAL fluid of cynomolgus monkeys (Fig. 6). Oral treatment of monkeys with Sch527123 (0.3 and 3 mg/kg) 2 h before the first bronchoscopy and lung lavage significantly attenuated the influx of neutrophils and total number of cells appearing in the second BAL sample (Fig. 6). The inhibition of the pulmonary neutrophilia by Sch527123 was 55 ± 19 and 84 ± 11% at oral doses of 0.3 and 3 mg/kg, respectively (ED50 = 0.3 mg/kg.).
Discussion
In the present report, the in vivo pharmacology of a novel, potent, and selective CXCR1 and CXCR2 antagonist, with significant oral bioavailability, was extensively evaluated in murine, rat, and primate models that replicate many of the important features of human COPD, such as pulmonary neutrophilia, goblet cell hyperplasia in the airway epithelium, and the secretion of airway mucus. These models include LPS and V2O5 challenge in rats and mice (Bonner et al., 2000; Takano and Nakagawa, 2001; Aoki et al., 2002) and repeat bronchoscopy and lung lavage in monkeys (Cohen and Batra, 1980; Krombach et al., 1994). In each of the animal models tested, Sch527123 significantly attenuated neutrophil recruitment and inhibited goblet cell hyperplasia or mucus production. These observations indicate that Sch527123 offers significant therapeutic potential for the treatment of inflammatory lung disorders in which neutrophilia and mucus hypersecretion mediate disease onset and development.
LPS challenge is often used to trigger an acute inflammatory response and intratracheal administration of LPS to rats and mice induces a rapid recruitment of neutrophils into the lungs (Spond et al., 2001; Aoki et al., 2002; Reutershan et al., 2005). This response is triggered by the release of neutrophil chemoattractants, such as cytokine-induced neutrophil chemoattractants and macrophage inflammatory protein-2, that are released into the lung tissue and circulation following LPS challenge (Takano and Nakagawa, 2001; Aoki et al., 2002; Mukaida, 2002). Based on the fact that the neutrophils of both mice and rats seem to lack a functional CXCR1 receptor (Cacalano et al., 1994) and the observation that Sch527123 does not bind to either rodent CXCR1 receptor (Fan et al., 2006; unpublished data), our studies strongly suggest that the inhibitory properties of Sch527123 reported here are due to CXCR2 receptor blockade alone. These results build upon a growing body of evidence suggesting that the CXCR2 receptor is a predominant receptor mediating neutrophil trafficking to the lung in several species (Qiu et al., 2003; Johnston et al., 2005; Stevenson et al., 2005; Thatcher et al., 2005).

Light micrograph of the lung sections of mice stained with PAS (A, B, and C) and H&E (D, E, F, and G). A, lung section from a saline (i.t.)—vehicle-treated group showing absence of any PAS+ cells in the airway epithelium. B, lung section from a LPS (i.t.)-vehicle-treated group showing a significant increase in the number of PAS+ cells. C, lung section from a LPS (i.t)-Sch527123 (3 mg/kg p.o.)-treated group showing an attenuation of PAS+ cells. D, lung section from saline (i.t.)-vehicletreated group showing well inflated alveoli with no neutrophilic infiltration. E, lung section from LPS (i.t.)-vehicle-treated group showing massive infiltration of neutrophils into the alveoli. F and G, lung section from LPS (i.t.)-Sch527123-treated animal showing moderate inhibition of neutrophilic recruitment. All sections are 50× magnification
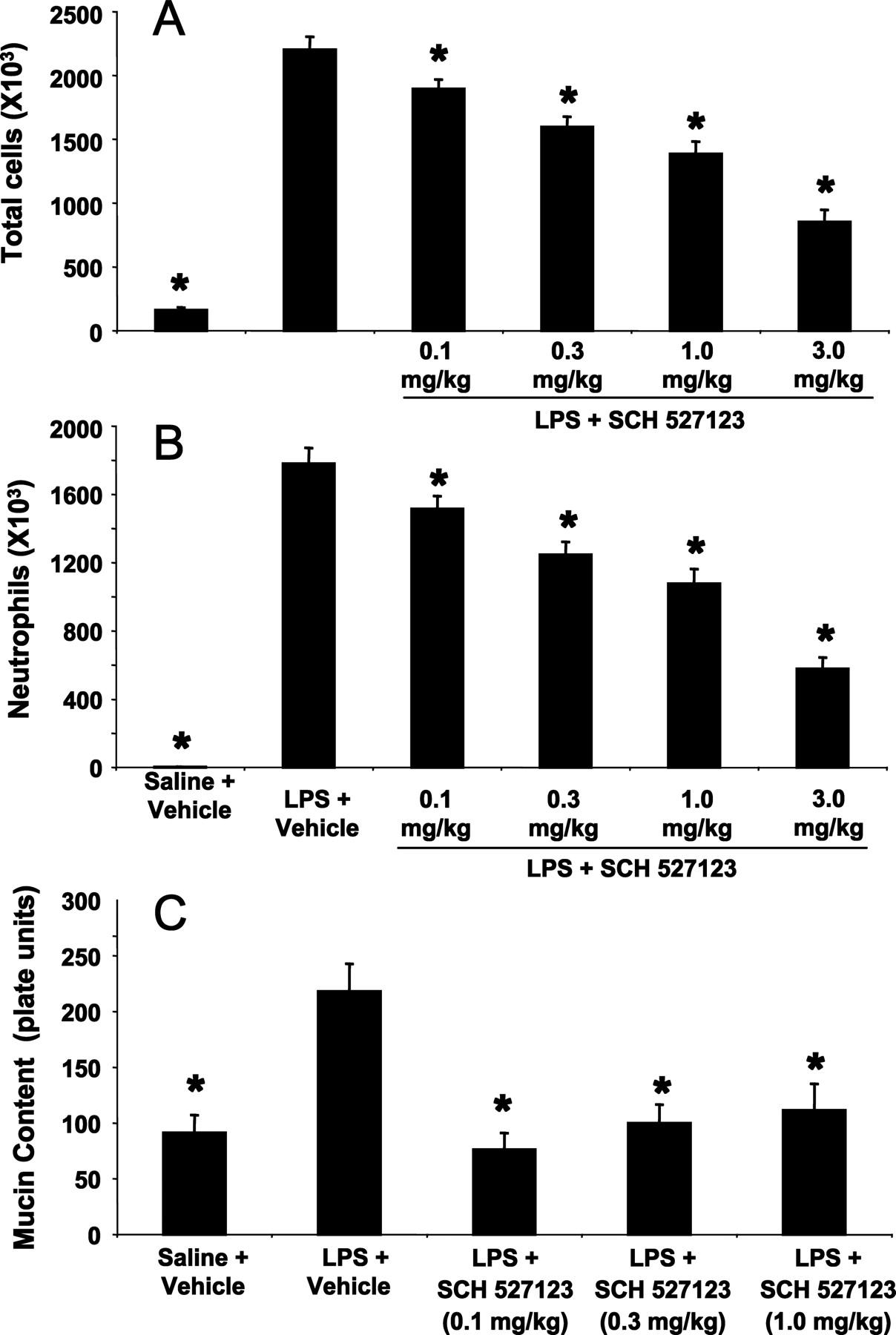
Sch527123 administration impairs neutrophil recruitment and mucus production in a rat model of intratracheal LPS challenge. A, total leukocytes in BALF. B, number of neutrophils in BALF. C, total mucin content. Data indicate the mean ± S.E.M. of 10 animals per treatment group. *, p < 0.05 compared to LPS + vehicle group.
Treatment with Sch527123 resulted in a sustained increase in the concentration of the CINCs in BAL fluid throughout 24 h after LPS challenge in rats. Similar results were obtained when Sch527123 was given to smoke-exposed mice (Thatcher et al., 2005). It is likely that CXCR2 blockade by Sch527123 alters the dynamics of ligand utilization in the context of the inflammatory response, leading to their elevation in these models. Of note, a different CXCR2 antagonist, SB-332235 [N-(2-hydroxy-3-sulfamyl-4-chlorophenyl)-N-(2,3-dichlorophenyl)urea], was found to inhibit CINC-2 production in lung tissue and BAL fluid of rats exposed to cigarette smoke (Stevenson et al., 2005). It is likely that differences in receptor affinity and binding kinetics between these compounds account for this discrepancy.

Sch527123 augments the production of CINC-2α and CINC-3 appearing in the BAL fluid of LPS-challenged rats. A BAL fluid sample was obtained 4 and 24 h after LPS. Data indicate mean ± S.E.M. of five animals per treatment group. *, p < 0.05 compared to LPS + vehicle group.
A significant histopathological feature of the LPS challenge in our study was the presence of goblet cell hyperplasia and an increase in BAL mucin. Pretreatment with Sch5127123 blocked the goblet cell hyperplasia and hypersecretion of mucus in these models, suggesting involvement of the CXCR2 receptors. Similar results were found when vanadium pentoxide, an airborne pollutant that has been associated with occupational asthma and bronchitis in humans (Levy et al., 1984; Bonner et al., 2000), was used as the inciting stimulus to induce the mucus phenotype. In contrast, Stevenson et al. (2005) found an inverse dose-response relationship for the inhibition of goblet cell density in smoke-exposed rats using SB-332235. As indicated above, dissimilar receptor affinity and/or binding kinetics between these compounds may contribute to these differences. In addition, it is important to note that we used relatively acute challenge models to evaluate the effects of Sch527123, whereas Stevenson et al. (2005) exposed rats to cigarette smoke for 3 consecutive days, implying that the chronicity of these different challenges may contribute to these disparities.

Sch527123 inhibits neutrophil recruitment, goblet cell hyperplasia, and mucus production in a rat model of intratracheal V2O5 challenge. A, number of neutrophils per milliliter of BALF. B, percentage of PAS-positive staining cells within the airway epithelium. C, mucin content. Data represent the mean ± S.E.M. of six animals per group. *, p < 0.05 compared to V2O5 + vehicle group.
Sch527123 was highly efficacious in impairing goblet cell hyperplasia and mucus secretion and in some models appeared to be more active in inhibiting the mucous phenotype than blocking neutrophil recruitment. These observations suggest that a modest attenuation of neutrophil recruitment to the lung may have profound protective effects on goblet cell biology, including mucus production. These findings are consistent with the concept that neutrophil elastase and reactive oxygen species directly contribute to goblet cell alterations in COPD patients (Kim and Nadel, 2004; Barnes and Stockley, 2005). Alternatively, direct CXCR2 inhibition on airway goblet cells may offer additional protection independent of the modulation of neutrophil trafficking. This latter possibility is consistent with the reported functional role of CXCR2 expressed on goblet cells (Smirnova et al., 2002; Miller et al., 2003).
Bronchoscopy and lung lavage produced a pulmonary neutrophilia in our cynomolgus monkeys, consistent with prior reports in nonhuman primates (Cohen and Batra, 1980; Krombach et al., 1994) and in humans (Chetta et al., 1992). This response is caused by airway closure and partial atelectasis due to the lavage procedure (Chetta et al., 1992). Furthermore, collapse and re-expansion of the lung triggers an inflammatory response that is characterized by neutrophilia and increased levels of CXCL8 in the lungs (Nakamura et al., 2000; Mukaida, 2002). The utility of this model is supported by our observation that CXCL8 levels in the BAL fluid increased from 56 ± 14 pg/ml at baseline to 438 ± 162 pg/ml at 6 h and 146 ± 88 pg/ml at 24 h. Therefore, repeat bronchoscopy and lavage produce an inflammatory response involving CXCL8 that most probably accounts for the elevated neutrophil recruitment to the lung. Interestingly, we were unable to reliably measure other ELR chemokines, such as CXCL1, from cynomolgus bronchoalveolar lavage fluid using human ELISA kits, probably because of poor cross-reactivity of the available human reagents.
In our cynomolgus monkey study, we found that oral administration of Sch527123 resulted in a significant, dose-dependent inhibition of neutrophil recruitment, with little apparent effect on the trafficking of other leukocyte cell types (Fig. 6). Based on the pharmacokinetic profile of Sch527123 in this species (Table 3) and its superior potency at the cynomolgus CXCR2 receptor relative to CXCR1 (Tables 1 and 2), we believe that the anti-inflammatory activity of Sch527123 in this model is mediated via CXCR2 antagonism, with a minimal contribution of CXCR1 blockade. Together with the results from our rodent models, these data support the concept that the CXCR2 receptor is the predominant chemokine receptor controlling neutrophil migration into the lungs under inflammatory conditions. This is a notable finding, given the broad array of receptors, which have been previously implicated in neutrophil trafficking in pulmonary diseases (Stockley, 2002; Barnes and Stockley, 2005). We interpreted the lack of marked efficacy of a humanized neutralizing anti-CXCL8 antibody administered to COPD patients (Mahler et al., 2004) as being consistent with this interpretation, because this approach would only partially inhibit the CXCR1/CXCR2 inflammatory axis. Interestingly, there was a significant positive association between neutrophils and CXCR2 but not CXCR1 mRNA expression in COPD patients with exacerbations (Qiu et al., 2003). This observation is consistent with a selective role of CXCR2 in mediating neutrophil trafficking to the lung under inflammatory conditions. Our data stand in contrast to previous reports in arthritis models in the rabbit, a species whose neutrophils also express both CXCR1 and CXCR2, which suggested that incomplete impairment of neutrophil recruitment is achieved by CXCR2-selective inhibition (Matsukawa et al., 1999; Podolin et al., 2002). Differences in the stimuli used and the site of inflammation may account for this discrepancy.

Treatment with Sch527123 inhibits neutrophil trafficking in a primate model of repeat bronchoscopy. A, total leukocytes per milliliter of BALF. B, number of neutrophils per milliliter of BALF. Hatched bars represent pre-bronchoscopy determinations. Filled bars represent post-bronchoscopy determinations. Data indicate the mean ± S.E.M. of six animals per treatment group. *, p < 0.05 compared to vehicle group.
The results obtained in our study have important implications for the role of CXCR2 receptors in chronic inflammatory lung disorders, such as COPD and asthma. We have demonstrated a significant and robust inhibition of pulmonary neutrophilia, goblet cell hyperplasia, and mucus hypersecretion with Sch527123. Despite being relatively transient in nature compared with the chronic decline in lung function in obstructive lung disorders in man, the animal models used for these evaluations demonstrate many of the same important features for both COPD and asthma, and the stimuli used may in fact contribute to the pathophysiology of these inflammatory lung disorders. For example, endotoxin is a significant and active component of cigarette smoke, which is the leading cause of COPD (Barnes and Stockley, 2005). Furthermore, patients with COPD often cycle through acute exacerbations, which frequently are triggered by infectious agents, including bacteria that may contain endotoxin on their cell wall (Stockley, 2002; Barnes and Stockley, 2005). Vanadium pentoxide is a metal contained in air pollution that may cause occupational asthma and bronchitis (Levy et al., 1984). Furthermore, the pulmonary inflammation induced by the repeat bronchoscopy procedure in primates is probably the result of regional airway closure and atelectasis (Cohen and Batra, 1980; Krombach et al., 1994), which is a condition that exists in obstructive lung disorders, such as COPD and asthma.
In summary, our observations suggest that Sch527123 would be of significant benefit in the treatment of human COPD and asthma, as well as other disorders that have pulmonary neutrophilia and mucus hypersecretion as an important part of their pathology, such as cystic fibrosis, acute lung injury, and acute respiratory distress syndrome. Furthermore, our data point to the central role of CXCR2 in mediating neutrophil recruitment and their sequelae during pulmonary inflammation in rodents, whose leukocytes selectively express CXCR2, and in primates, whose neutrophils express both CXCR1 and CXCR2. We speculate that a component of the anti-inflammatory activity of Sch527123 may also be mediated via endothelial and epithelial cells, because CXCR2 expression on these cell types has been implicated in neutrophil recruitment and lung injury (Reutershan et al., 2006). Our ongoing efforts continue to focus on further characterizing the therapeutic profile of Sch527123 in both acute and chronic inflammatory conditions.
Acknowledgments
We gratefully acknowledge Howard Jones, Yvette Crawley, Jennifer Richard, Natalie Case, Susan Sehring, Xiang Y. Zhang, Susan Skeans, and James Lamca for superb technical assistance. We also thank Drs. Arthur Taveras, Robert Egan, Satwant Narula, and Daniel Lundell for advice and support.
Footnotes
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.106.119040.
-
ABBREVIATIONS: COPD, chronic obstructive pulmonary disease; BAL, bronchoalveolar lavage; CINC, cytokine-induced neutrophil chemoattractant; Sch527123, 2-hydroxy-N,N-dimethyl-3-[[2-[[1(R)-(5-methyl-2-furanyl)propyl]amino]-3,4-dioxo-1-cyclobuten-1-yl]amino]benzamide; Sch-N, Sch527123; CXCL1, growth-related protein-α; CXCL5, epithelial-derived neutrophil attractant-78; CXCL7, neutrophil-activating peptide-2; CXCL8, interleukin-8; ELR, glutamine-leucine-arginine motif; i.t., intratracheal; LPS, lipopolysaccharide; MIP-2, macrophage inflammatory protein-2; PAS, periodic acid Schiff; V2O5, vanadium pentoxide; H&E, hematoxylin and eosin; ELISA, enzyme-linked immunosorbent assay; SB-332235, N-(2-hydroxy-3-sulfamyl-4-chlorophenyl)-N-(2,3-dichlorophenyl)urea.
- Received December 21, 2006.
- Accepted April 25, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}