


Abstract
Functional studies have demonstrated that adrenoceptor agonist-evoked relaxation is mediated primarily by β3-adrenergic receptors (ARs) in human bladder. Thus, the use of selective β3-AR agonists in the pharmacological treatment of overactive bladder is being explored. The present studies investigated the effects of a novel selective β3-AR agonist, (R)-3′-[[2-[[2-(3-chlorophenyl)-2-hydroxyethyl]amino]ethyl]amino]-[1,1′-biphenyl]-3-carboxylic acid (GW427353; solabegron) on bladder function in the dog using in vitro and in vivo techniques. GW427353 stimulated cAMP accumulation in Chinese hamster ovary cells expressing the human β3-AR, with an EC50 value of 22 ± 6 nM and an intrinsic activity 90% of isoproterenol. At concentrations of 10,000 nM, GW427353 produced a minimal response in cells expressing either β1-ARs or β2-ARs (maximum response <10% of that to isoproterenol). In dog isolated bladder strips, GW427353 evoked relaxation that was attenuated by the nonselective β-AR antagonist bupranolol and 1-(2-ethylphenoxy)-3-[[(1S)-1,2,3,4-tetrahydro-1-naphthalenyl]amino]-(2S)-2-propanol (SR59230A) (reported to have β3-AR antagonist activity). The relaxation was unaffected by atenolol, a selective β1-AR antagonist, or (±)-1-[2,3-(dihydro-7-methyl-1H-inden-4-yl)oxy]-3-[(1-methylethyl)amino]-2-butanol (ICI 118551), a selective β2-AR antagonist. GW427353 increased the volume required to evoke micturition in the anesthetized dog following acetic acid-evoked bladder irritation, without affecting the ability of the bladder to void. GW427353-evoked effects on bladder parameters in vivo were inhibited by bupranolol. The present study demonstrates that selective activation of β3-AR with GW427353 evokes bladder relaxation and facilitates bladder storage mechanisms in the dog.
Overactive bladder is a syndrome characterized by symptoms of urinary frequency, urgency, nocturia, and urgency incontinence in which there is a need for alternative therapies with novel mechanisms of action (Andersson and Wein, 2004). The activities of the urinary bladder during urine storage and voiding are governed by a complex neural control system, involving both sympathetic and parasympathetic input. During bladder filling, sympathetic nerve activity to bladder smooth muscle results in β-adrenergic receptor (AR)-mediated relaxation (Andersson and Wein, 2004). Several subtypes of β-ARs have been identified, namely, β1, β2, and β3-ARs (Bylund et al., 1994). Evidence suggests that species differences exist in the distributions and roles of β-ARs in the bladder (for comprehensive review, see Michel and Vrydag, 2006). Bladder relaxation evoked by β-AR agonists is mediated mainly via β1-ARs in cats (Nergardh et al., 1977) and guinea pigs (Li et al., 1992), by β2-ARs in rabbits, by both β2- and β3-ARs in rats (Yamazaki et al., 1998) and pigs (Yamanishi et al., 2002), and by β3-ARs in the ferrets (Takeda et al., 2000), dogs (Yamazaki et al., 1998), and primates (Takeda et al., 2002a). Based on mRNA expression, at least 95% of the adrenoceptor message in human bladder comprises the β3-AR subtype (Nomiya and Yamaguchi, 2003). Furthermore, functional studies have demonstrated that the relaxant effects induced by adrenergic receptor stimulation in human detrusor are mediated via β3-AR activation (Yamaguchi, 2002; Yamanishi et al., 2006). Thus, selective β3-AR stimulation of human bladder would be expected to cause bladder relaxation, and as such it may represent a novel strategy in the treatment of overactive bladder. The present studies investigated the effects of a novel, selective β3-AR agonist, GW427353 (solabegron; Uehling et al., 2006), on bladder function in the dog using in vitro and in vivo techniques.
Materials and Methods
All experiments were reviewed and approved by the Institutional Animal Care and Use Committee of GlaxoSmithKline. All procedures on animals were performed in compliance with Institute of Laboratory Animal Resources (1996).
Radioligand Binding Assay
Membranes from Chinese hamster ovary (CHO) cells expressing β1-, β2-, or β3-ARs were prepared as described previously (Elshourbagy et al., 1993). Saturation binding experiments were performed as described previously (Schnabel et al., 2000), except a 50-μl volume was used and incubation was for 30 min at 37°C. A range of final concentrations of [125I](–)-iodocyanopindolol (ICYP), 10 to 600 pM for β1 and β2 and 0.1 nM to 2.3 nM for β3 were used. Nonspecific binding was measured in the presence of 10 μM SR59230A. Protein was measured with a Bio-Rad protein assay kit (Bio-Rad, Hercules, CA). Receptor affinity and density for the various receptors were calculated by global fit of total and nonspecific binding curves using GraphPad Prism version 4.0 (GraphPad Software Inc., San Diego, CA).
Measurement of Cyclic AMP
CHO-K1 cells stably expressing β-ARs were grown in 50:50 Dulbecco's modified Eagle's medium plus F-12 K with Geneticin (G-418; Invitrogen, Carlsbad, CA) and hygromycin in 24-well plates (50,000 cells/well) for 48 h. On the day of experiment, medium was replaced with phosphate-buffered saline (Invitrogen) containing 1 mM 3-isobutyl-1-methylxanthine. Cells were then treated with vehicle (2.5% DMSO), GW427353 in 2.5% DMSO, or isoproterenol in DMSO (40× concentration) in 200 μl of phosphate-buffered saline containing 1 mM 3-isobutyl-1-methylxanthine and incubated at 37°C for 30 min. After incubation cells, were treated with lysis buffer, and cAMP level was determined using the commercial kit Amersham SPA as per package instructions of the manufacturer (GE Healthcare, Little Chalfont, Buckinghamshire, UK). The SPA measures cAMP via competition between cAMP and adenosine 3′,5′-cyclic phosphoric acid 2′-O-succinyl-3-[125I]iodotyrosine methyl ester for an antibody/scintillant microbead system. Samples were processed under the nonacetylation protocols for the kit. The range of sensitivity for the nonacetylation protocols of the kit was 0.2 to 12.8 pmol/assay tube. Aliquots (50 μl) of the samples were transferred to 96-well OptiPlate (PerkinElmer Life and Analytical Sciences, Boston, MA) for determining cAMP level. The plates were counted on a TopCount scintillation counter (PerkinElmer Life and Analytical Sciences, Boston, MA). All experiments were performed in duplicate, with n referring to the number of independent experiments. Values are expressed as mean ± S.E.M. throughout the article.
In Vitro Urinary Bladder Tissue Assay
Mongrel and beagle dogs (9–12 kg; Marshall Farms, North Rose, NY) were euthanized with an injection of 390 mg/ml sodium pentobarbital into the cephalic vein followed by rapid exsanguination. The bladder was exposed and removed, fat and connective tissue were cleared, and the bladder body was cut into longitudinal strips approximately 3 × 6 mm. The strips were suspended in tissue baths containing Krebs' solution (118 mM NaCl, 4.7 mM KCl, 2.5 mM CaCl2, 1.5 mM MgSO4, 1.2 mM K2HPO4, 25 mM NaCO3, and 11 mM glucose). Tissues were maintained at 37°C, and they were constantly aerated with 95% oxygen and 5% CO2. Tissues were mounted to a force-displacement transducer (Biopac Systems, Inc., Goleta, CA), and changes in muscle tension were digitally recorded using Acq-Knowledge 3.8.1 software (Biopac Systems, Inc.).
Tissues were equilibrated for 60 min, during which the tissues were maintained at 9.8 mN resting tension. Concentration-response curves for GW427353 were obtained by sequential addition (30-min interval between doses) directly into the bath 20 min after stimulation with 15 mM KCl. Papaverine (100 μM) was added following the construction of concentration-response curves. In a separate group of experiments, the effects of β-AR antagonists (bupranolol, atenolol, ICI 118551, and SR59230A) were assessed on a single dose of 10 nM GW427353. Antagonist or vehicle was added 5 min after 15 mM KCl stimulation, and tissue was left to incubate for 15 min before addition of GW427353. After a 30-min incubation with GW427353, 100 μM papaverine was added. Stock solutions were prepared as follows: papaverine, ICI 118551, and SR59230A were dissolved in water, GW427353 and bupranolol were dissolved in DMSO, and atenolol was dissolved in ethanol, and the solutions were diluted to desired concentrations in water. The reported concentrations are the calculated final bath concentrations.
Data Analysis. Integrals over a 2-min period were measured before and after addition of test substances. The effects of GW427353 were expressed as percentages of the maximal relaxation induced by 100 μM papaverine. In cumulative concentration-response experiments, GW427353-evoked responses were compared with vehicle-treated tissues using two-way analysis of variance followed by Bonferroni's post-test. In single-concentration experiments, GW427353-evoked responses in the absence or presence of antagonist were compared using unpaired t test. P < 0.05 was considered statistically significant.
In Vivo Acetic Acid-Evoked Bladder Irritation in the Dog
Experiments were performed on 29 adult female beagle dogs (6–10 kg; Marshall Farms) fasted overnight but given free access to water. Cannulae were inserted into both cephalic veins for intravenous (i.v.) administration of anesthetic and test substances. Animals were then administered 6 mg/kg i.v. propofol for anesthetic induction. Anesthesia was maintained with propofol infusion (5 mg/min i.v.). The trachea was intubated to maintain a patent airway. The left femoral artery was cannulated with a heparinized cannula [20 U ml–1 heparin in 0.9% (w/v) saline; polyethylene 50] for the measurement of mean arterial blood pressure (MAP) and heart rate (HR) and for sampling arterial blood for blood gas analysis. Body temperature was maintained between 36 and 38°C using a blanket system (TP-500; Gaymar Industries, Inc., Orchard Park, NY). Blood pressure was measured using a pressure transducer (P23XL; Gould Statham, Oxnard, CA), and the HR was derived electronically online from the blood pressure signal using Spike version 5.0 software (Cambridge Electronic Design, Cambridge, UK). The urinary bladder was exposed by a midline abdominal incision, and a cut was made in the bladder dome and a cannula (polyethylene 60) was inserted into the bladder via a 14-gauge needle. The needle was removed, and the cannula was secured with a purse string suture. The cannula, via a three-way tube connector (Small Parts Inc., Miami Lakes, FL), was connected to a pressure transducer (P23XL; Gould Statham) to record intravesical bladder pressure and to a syringe pump (model 975 infusion pump; Harvard Apparatus Inc., Holliston, MA) for the intravesical infusion of saline or acetic acid. Draining through the cannula allowed the bladder to be emptied of saline. Void volume was measured by collecting and weighing fluid released from the bladder during micturition. At the end of all surgical procedures, propofol anesthesia was discontinued, and 100 mg/kg i.v. α-chloralose was administered. Depth of anesthesia was assessed by the stability of MAP and HR and by an absence of hind limb withdrawal in response to paw pinch. Anesthesia was maintained via 1 mg/kg/min i.v. α-chloralose infusion. Animals were artificially ventilated with room air by use of a positive pressure pump (Large Animal Ventilator, model 613; Harvard Apparatus Inc.). Blood gases were monitored and maintained between 90 and 130 mm Hg pO2, between 20 and 35 mm Hg pCO2, and between pH 7.3 and 7.4 with a pH/blood gas analyzer (model 248; Corning Life Sciences, Acton, MA). Adjustments of the respiratory pump rate and volume were made as necessary to maintain blood gas and pH balance. After completion of studies, dogs were euthanized with 120 mg/kg i.v. sodium pentobarbital without recovery.
After surgery, animals were allowed to stabilize for 30 min during which blood gases were monitored and maintained. Warmed saline [0.9% (w/v)] was then infused into the bladder at 3 ml/min until voiding (the micturition reflex) occurred, characterized by a large-amplitude bladder contraction with the concomitant release of fluid from the urethral opening. This fluid was measured and termed the void volume (milliliters). Micturition volume threshold (milliliters) was taken as the total volume of fluid required to be infused into the bladder to evoke voiding. Voiding efficiency (percentage) was calculated from voided volume divided by micturition volume threshold and multiplied by 100. Fluid was emptied from the bladder following voiding for a 10-min period (until the bladder was as empty as possible), after which the bladder was reinfused with saline to evoke a further micturition reflex. This was continued until four control voids had been evoked. GW427353 or vehicle (polyethylene glycol 400) was then administered as a bolus i.v. at 0.15 ml/kg followed by a 5-min period to allow stabilization of any change in baseline parameters. The saline infusion into the bladder was then replaced with 0.7% acetic acid in 0.9% saline, pH 3.0, and three acetic acid voids were evoked as outlined in the protocol described above. This time period was chosen because preliminary studies showed a reproducible hyperactive bladder response to acetic acid could be evoked using this protocol. Blood samples (200 μl) were taken following each void for plasma sample analysis of drug levels.
Data Analysis. Arterial blood and bladder pressures were continuously displayed on a chart recorder (Grass Instruments, Quincy, MA) and captured (200 samples/s) by a CED 1401 interface (Cambridge Electronic Design) to allow data to be acquired and analyzed off-line using Spike version 5 software (Cambridge Electronic Design). HRs were derived electronically online from the blood pressure signal using this software. All baseline values were the mean values over a 30-s period, and they were measured 1 min before infusion of saline or acetic acid into the bladder. The amplitude (mm Hg) and duration (seconds) of each reflex-evoked bladder contraction was measured. The micturition pressure thresholds were taken as the bladder pressure (mm Hg) at which voiding occurred. The control values for various parameters were calculated from the mean of the last two preacetic acid voids (micturition parameters) or the mean of the last three voids (MAP and HR). The mean of the next three voids after treatment with vehicle or GW427353 was used to measure the change in void parameters, MAP, and HR. Changes were compared with control values and treatment values using a paired t or Student's t test as appropriate.
Plasma Analysis
Analysis of female dog plasma samples for GW427353 was performed using liquid chromatography/tandem mass spectrometry. Analytical standards for GW427353 were prepared in 50 μl of female dog plasma. Dog plasma samples were thawed, and plasma proteins were precipitated with 200 μl of acetonitrile/10 mM aqueous ammonium formate (95:5; pH 3.0), containing 200 ng/ml of a mass spectral internal standard. The resulting mixture was vortex-mixed for 2 min followed by centrifugation for 30 min at >2000g. Then, 2.0 μlofthe resulting supernatant was injected onto the liquid chromatography/tandem mass spectrometry system using an HTS PAL autosampler (CTC Analytics, Zwingen, Switzerland). The mobile phase consisted of a 2.1-min linear gradient using aqueous 10 mM ammonium formate, pH 3.0, and acetonitrile mobile phases (500 μl/min flow rate). A Polaris C8 Ether analytical column (2 × 50 mm, 3 μm; Varian, Inc., Lake Forest, CA) was used. The eluent flowed into a Sciex API4000 triple-quadrupole mass spectrometer (Applied Biosystems, Foster City, CA) using positive-ion electrospray multiple-reaction monitoring. GW427353 was characterized by the transition of the m/z 411 parent (M + H)+ precursor ion to its m/z 240 product ion, generated at an optimized collision energy.
Data Analysis. Data were reported as quantitative concentrations as determined by standard calibration curve analysis, using quadratic fitting of a 1/x-weighted plot of the GW427353/mass spectral internal standard peak area ratios versus GW427353 concentration, where the analytical range of quantitation was 50 to 10,000 ng/ml (r = 0.9944), lower limit of quantification to upper limit of quantification, respectively.
Plasma Protein Binding
Two male beagle dogs (8.5 and 14.5 kg) were fasted overnight but given free access to water. Cannulas were inserted into both cephalic veins, and animals were dosed with GW427353 i.v. (over a 5-min infusion period, in 0.025 M methanesulfonic acid in 5% mannitol) with a total dose of 0.2 mg/kg via one of the catheters. Five minutes after dosing, 2-ml blood samples were collected via the other cephalic vein. The plasma was separated from red blood cells by centrifugation, and extracts were frozen at –20°C. The extracts were analyzed for GW427353 using similar analytical methodologies as described above. Protein plasma binding was determined by ultrafiltration at 37°C using an Amicon Centrifree disposable ultrafiltration device (Millipore Corporation, Bedford, MA). An absorption test indicated that GW427353 did not bind to the membrane or to plastic components of the ultrafiltration device.
Materials
GW427353 (Uheling et al., 2006) and 1-(2-chloro-5-methylphenoxy)-3-[(1,1-dimethylethyl)amino]-2-propanol (bupranolol hydrochloride) were synthesized at GlaxoSmithKline Pharmaceuticals (King of Prussia, PA). Potassium chloride, papaverine hydrochloride, and (±)-4-[2-hydroxy-3-[(1-methylethyl)amino]propoxy]benzeneacetamide (atenolol) were purchased from Sigma-Aldrich (St. Louis, MO). ICI 118551 hydrochloride and SR59230A hydrochloride were purchased from Tocris Cookson Inc. (Bristol, UK). [125I](–)-Iodocyanopindolol (2200 Ci/mmol) was purchased from PerkinElmer Life and Analytical Sciences.
Results
In Vitro Cellular Assays
GW427353 stimulated cAMP accumulation in CHO cells expressing the recombinant human β3-AR, with an EC50 value of 22 ± 6.0 nM, and it produced a maximum response 90 ± 4% of that produced by the nonselective β-adrenoceptor agonist isoproterenol. At concentrations of 10,000 nM, GW427353 produced a minimal response in cells expressing either β1-ARs or β2-ARs (maximal response <10% of that to isoproterenol) (Table 1). GW427353 did not produce antagonism at any of the β-ARs (data not shown). Receptor density was determined in membranes from CHO cells expressing the recombinant β-ARs. β3-AR expression was found to be lower than that of both β1- and β2-AR expression (Table 2).
Effect of isoproterenol and GW427353 on cAMP accumulation in CHO cells expressing human recombinant β-adrenoceptors
Values are mean ± S.E.M.
Relative receptor density in CHO cell membranes expressing recombinant β-ARs and affinity for ICYP
Values are mean ± S.E.M.
In Vitro Urinary Bladder Tissue Assay
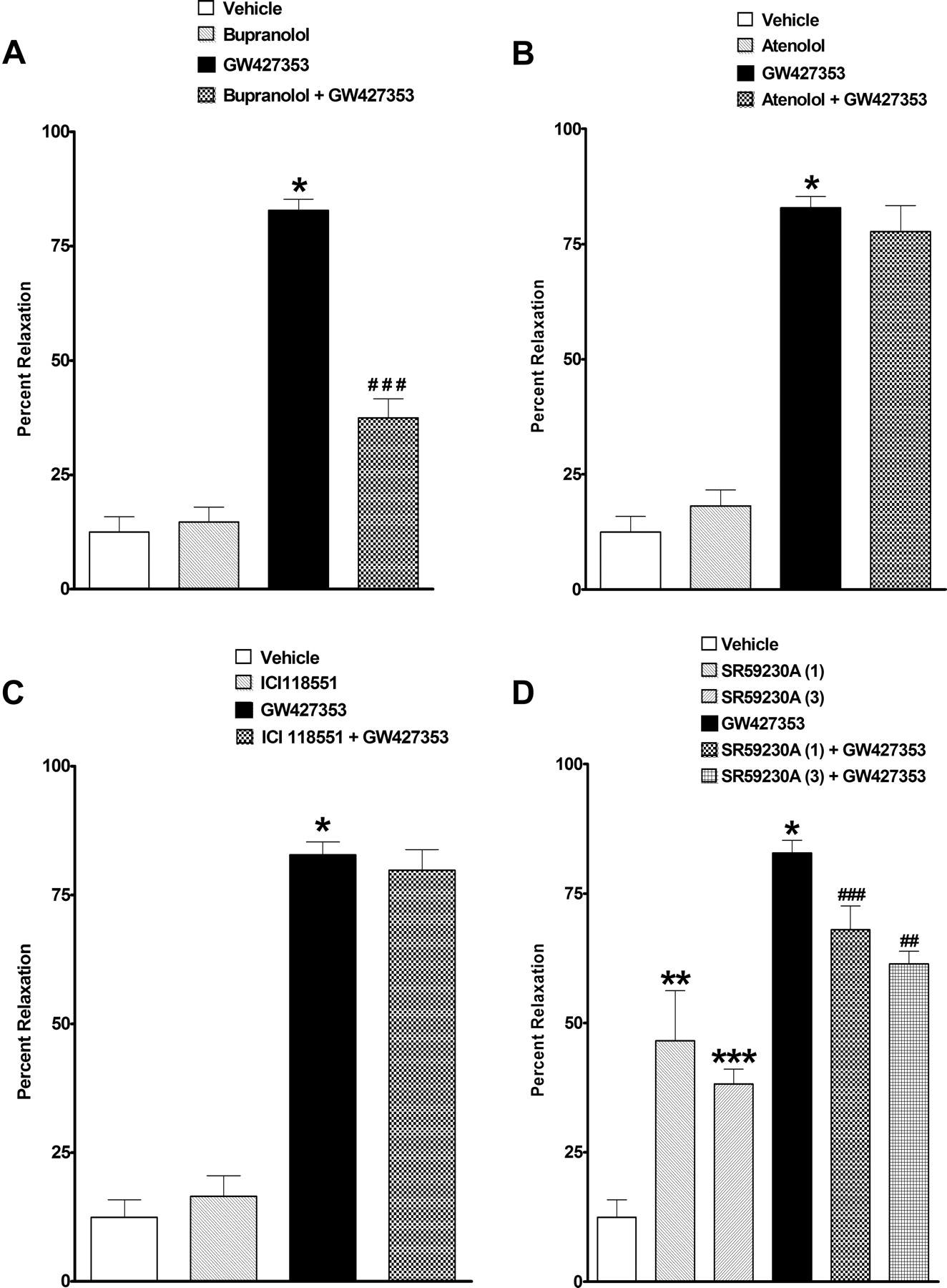
Cumulative addition of GW427353 evoked concentration-dependent relaxation of KCl precontracted dog bladder strips, attaining a significant effect at 100 nM (n = 18; Fig. 1). However, significant decreases in the tension of strips was observed over time in vehicle-treated tissues; thus, a single-concentration experimental protocol was used to evaluate the effects of antagonists. In the single-concentration experiments, decreases in the presence of vehicle were much less (12 ± 3.3%). A single concentration of 10 nM GW427353 produced 82 ± 2.5% relaxation (n = 6; Fig. 2A). Higher concentrations of GW427353 (30 and 100 nM) produced a degree of relaxation that was not statistically different from 10 nM (77 ± 6.9 and 89 ± 4.7%, respectively; n = 8; data not shown); thus, 10 nM GW427353 was used as the agonist concentration in antagonist studies. The nonselective β-AR antagonist bupranolol at 10 μM significantly attenuated the relaxation evoked by 10 nM GW427353 by approximately 64%, without having significant activity alone (n = 4; Fig. 2A). Neither the selective β1-AR antagonist atenolol at 1 μM nor the selective β2-AR antagonist ICI 118551 a 1 μM had any effects of bladder responses evoked by GW427353 (n = 8; Fig. 2, B and C). SR59230A, reported to have antagonistic properties at the β3-AR (Manara et al., 1996), evoked a significant relaxation of isolated canine bladder strips alone, evoking maximal responses of 47 ± 17 and 38 ± 5.8% at 1 and 3 μM, respectively (n = 4; Fig. 2D). In the presence of 1 and 3 μM SR59230A, GW427353-evoked relaxations were significantly attenuated by 21 and 30%, respectively (n = 4; Fig. 2D).

Effect of GW427353 on KCl-precontracted canine bladder strips. *, P < 0.05; **, P < 0.01 compared with vehicle-treated tissues.
In Vivo Acetic Acid-Evoked Bladder Irritation in the Dog
Control Micturition Reflexes. Infusion of saline into the bladder evoked the micturition reflex, characterized by the appearance of a large-amplitude (56 ± 5.8 mm Hg; n = 22) bladder contraction lasting 34 ± 7.2 s (n = 23), with the concomitant release of 27 ± 5.3 ml of fluid from the urethral opening (void volume; n = 23). The mean volume infused to evoke the micturition reflex (micturition volume threshold) was 38 ± 4.3 ml (n = 23) with a calculated voiding efficiency (voided volume as a percentage of micturition reflex threshold) of 84 ± 13% (n = 23). This parameter was sometimes calculated to be greater than 100%, because not all the residual volume could be drained from the bladder before the start of the next bladder infusion. Manual expression of residual urine from the bladder was not performed to prevent potential sensitization of bladder nociceptive afferent nerves during the control period. The mean bladder pressure threshold to evoke the micturition reflex was 38 ± 3.9 mm Hg (micturition pressure threshold; n = 22). Baseline MAP and HR were 123 ± 5.6 mm Hg and 116 ± 7.3 beats per min, respectively (n = 22). The mean baseline data for bladder and cardiovascular parameters are shown in Table 3.
Summary of baseline and treatment data for bladder and cardiovascular parameters in dogs treated with vehicle (n = 7) or GW427353 (1 mg/kg, n = 3; 3 mg/kg, n = 7)
Baseline values represent the mean of the last two voids before initiation of acetic acid infusion. Vehicle and GW427353 values represent the mean of three voids following infusion of acetic acid. Values are mean ± S.E.M.
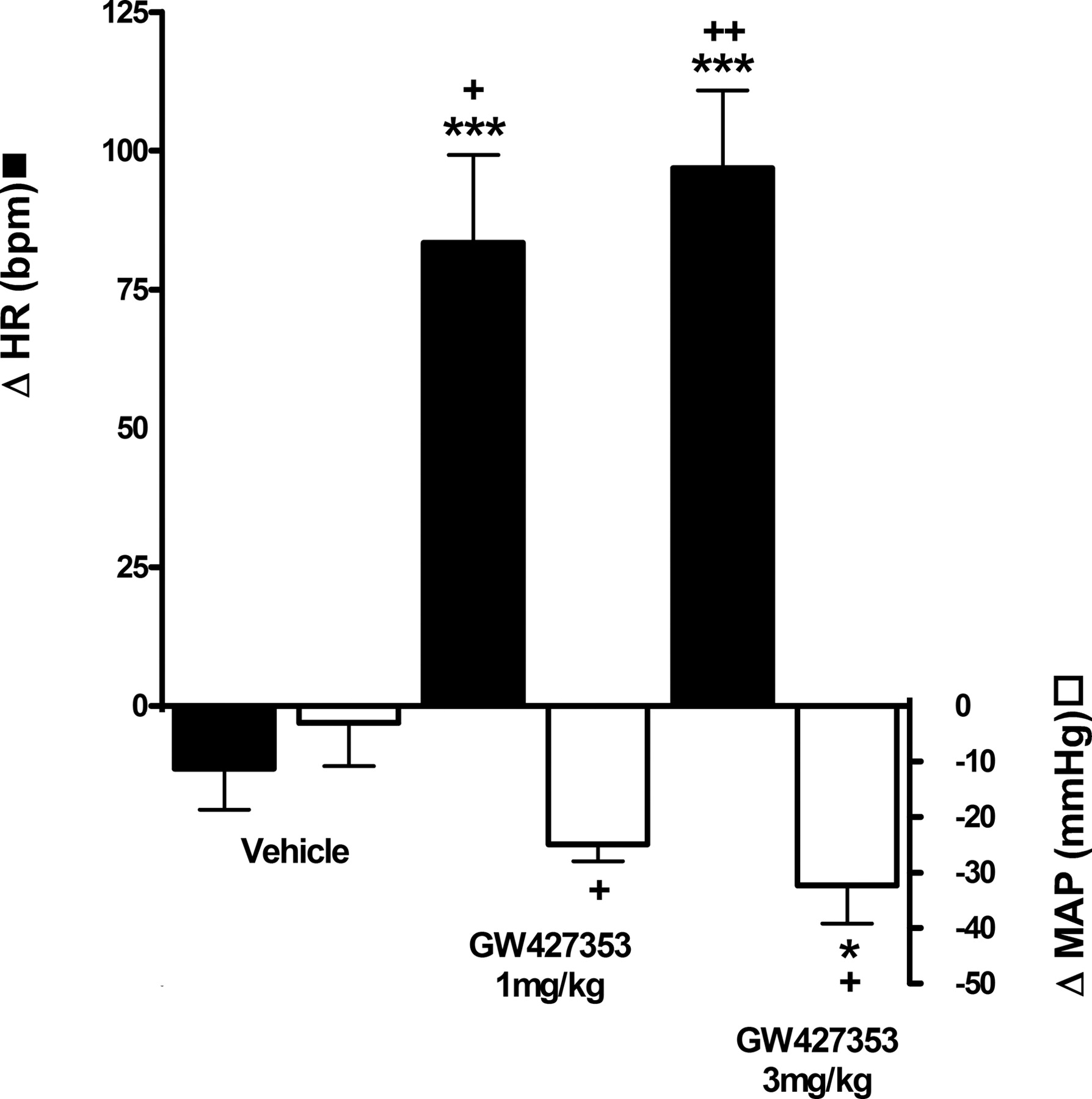
Effects of GW427353 on Acetic Acid-Evoked Bladder Irritation. In vehicle-pretreated dogs, intravesical infusion of acetic acid evoked a decrease in the micturition volume threshold of 10 ± 2.3 ml or 29% (n = 7) (Table 3; Fig. 3). Intravesical acetic acid facilitated voiding, evoking a concomitant increase in void volume, voiding efficiency, and bladder contraction amplitude, probably as a result of residual volumes that were not emptied during the control period now being evacuated (Table 3). Acetic acid had no effect on micturition pressure threshold, bladder contraction duration, MAP, and HR (Table 3). GW427353 at 3 mg/kg i.v. evoked an increase in micturition volume threshold of 6.5 ± 4.4 ml or 22%; thus, it prevented the acetic acid-evoked decreases in this parameter (Table 3; Fig. 3). A lower dose of GW427353, 1 mg/kg i.v., seemed to block the reduction in micturition threshold produced by acetic acid, but this effect was not significant (Fig. 3). Neither dose of GW427353 had any effect on void volume, voiding efficiency and bladder contraction amplitude, and duration (Table 3). Both doses of GW427353 evoked a significant decrease in MAP (25 ± 3.0 mm Hg or 22% and 35 ± 5.3 mm Hg or 30%) and increase in HR (83 ± 16 mm Hg or 105% and 97 ± 14 mm Hg or 82%) after administration of 1 and 3 mg/kg, respectively (Table 3; Fig. 4). Flushing of the dogs was also observed, but it was not measured.
In vivo antagonist experiments were performed with the higher dose of GW427353, 3 mg/kg i.v. Bupranolol at 1 mg/kg i.v. alone had no significant effects on micturition threshold volume (Fig. 5) or cardiovascular parameters compared with vehicle treatment (data not shown). Pretreatment with bupranolol prevented the GW427353-evoked increases in micturition threshold volume (Fig. 5). In addition, bupranolol significantly attenuated the GW427353-evoked increase in HR but not the GW427353-evoked decrease in MAP (Fig. 6).

Effect of 10 nM GW427353 on KCl-precontracted bladder strips alone and in the presence of 10 μM bupranolol (A), 1 μM atenolol (B), 1 μM ICI 118551 (C), and 1 and 3 μM SR59230A (D). *, P < 0.05; **, P < 0.01; ***, P < 0.001 for comparison of effects of compound versus vehicle. ##, P < 0.01; ###, P < 0.001 for comparison of GW427353 effect in the presence and absence of antagonist.

Effect of GW427353 on micturition threshold in the anesthetized dog treated with intravesical acetic acid. Each bar represents the mean of seven (vehicle and 3 mg/kg GW427353) or three (1 mg/kg GW427353) experiments ± S.E.M. Significant difference: ++, P < 0.01 from pretreatment; and **, P < 0.01 from vehicle.
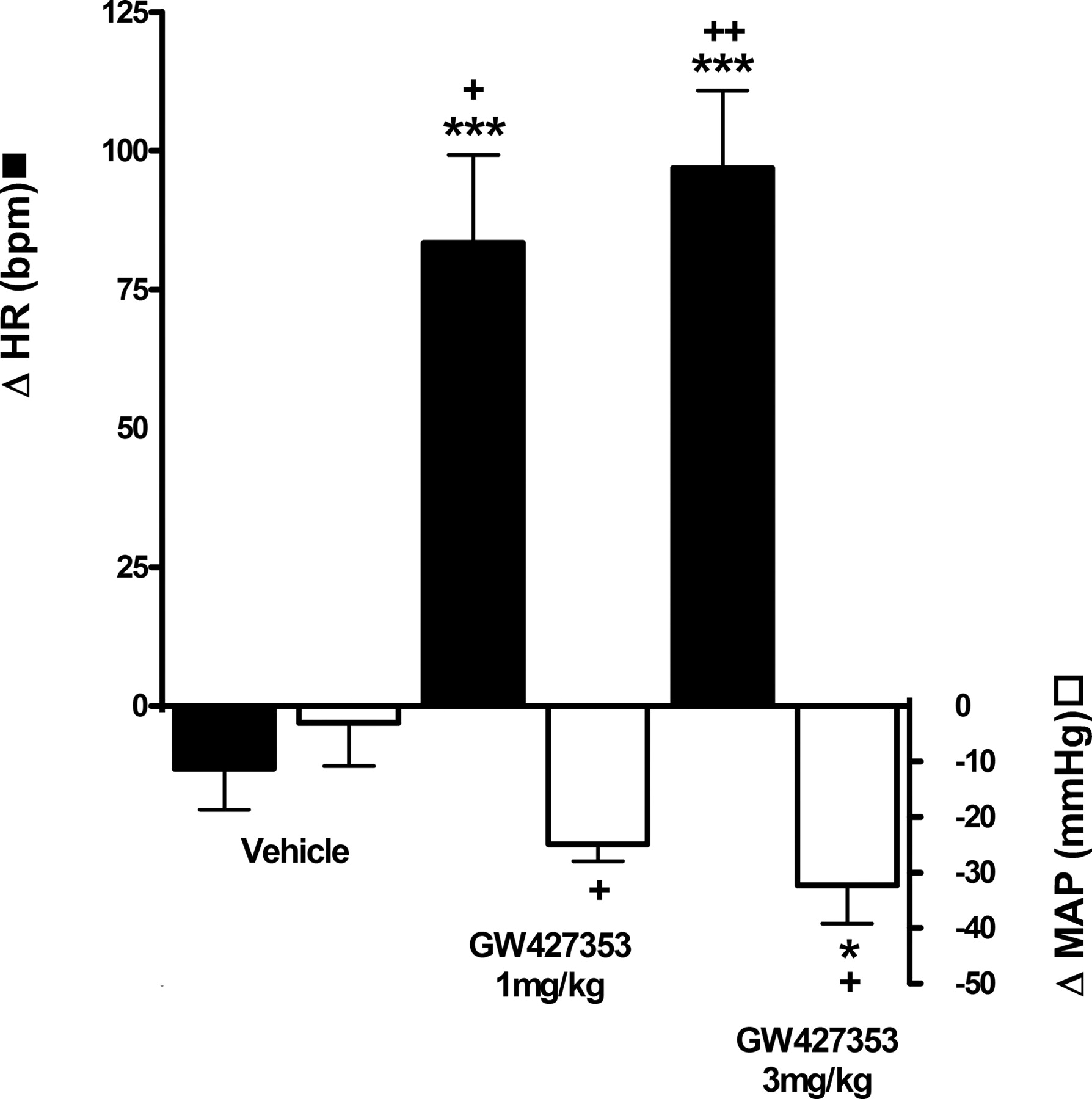
Effect of GW427353 on HR and MAP in the anesthetized dog. Values represent the mean of six (vehicle and 3 mg/kg GW427353) or three (1 mg/kg GW427353) experiments ± S.E.M. Significant difference: +, P < 0.05; ++, P < 0.01 from predosing; *, P < 0.05 and ***, P < 0.001 from vehicle.
Plasma Analysis and Protein Binding
Plasma samples were taken after each acetic acid void following i.v. dosing with GW427353 in the above-mentioned experiments, and plasma concentrations were determined (Table 4). Average plasma concentrations were 2130 ng/ml (4.8 μM) and 3343 ng/ml (7.5 μM) after doses of 1 and 3 mg/kg, respectively. In separate experiments, dog plasma protein binding of GW427353 was determined to be 99.3% (Table 5).
Plasma levels of GW427353 attained following each void after initiation of acetic acid infusion in the dog following i.v. administration Values are mean ± S.E.M. in nanograms per milliliter. Values in parentheses are in micromolar.
Protein binding of GW427353 in the dog following i.v. administration (0.2 mg/kg)

Effects of intravenous administration of GW427353, bupranolo1, and the combination of bupranolol and GW427353 on change in micturition threshold in anesthetized dog. Bars represent the mean of six to seven (vehicle and GW427353) or three (bupranolol and bupranolol + GW427353) experiments ± S.E.M. Significant difference: *, P < 0.05 from vehicle; and ##, P < 0.01 from GW427353.

Effects of intravenous administration of GW427353, bupranolo1, and the combination of bupranolol and GW427353 on change in cardiovascular parameters in anesthetized dog. Bars represent the mean of seven (GW427353) or three (bupranolol and bupranolol + GW427353) experiments ± S.E.M. Significant difference: #, P < 0.05 from GW427353.
Discussion
The present study demonstrates that stimulation of β3-ARs with GW427353 evokes bladder relaxation and facilitates bladder storage mechanisms in the dog, suggesting that β3-AR agonists may represent a novel approach in the treatment of overactive bladder. In human bladder, at least 95% of the adrenoceptor message comprises the β3-AR subtype, and adrenoceptor agonist-evoked relaxation is mediated principally by these receptors (Yamaguchi, 2002; Nomiya and Yamaguchi, 2003; Yamanishi et al., 2006). GW427353 has been found to relax isolated human bladder strips and to reduce spontaneous contractile activity (Biers et al., 2006; T. D. Westfall, unpublished observations). In rat models, β3-AR agonists have been shown to increase micturition interval in bladder instability evoked by cerebral infarction (Kaidoh et al., 2002), intravesical acetic acid or prostaglandin E2 (Woods et al., 2001; Takeda et al., 2002b); inhibit spontaneous intermicturition contractions in obstruction-induced bladder hyperplasia (Woods et al., 2001); and decrease the frequency of rhythmic bladder contractions (Takasu et al., 2007). β3-AR-mediated bladder relaxation is also observed in the anesthetized ferret (Takeda et al., 2000).
GW427353 is a highly selective β3-AR agonist as evidenced by selective stimulation of cAMP accumulation in CHO cells expressing the human β3-AR, with an intrinsic activity nearly equivalent to that of isoproterenol. GW427353 did not possess antagonistic activity at any of the β-ARs (binding experiments with GW427353 were not included, because they would not add to the functional data). In an in vitro functional bladder assay, GW427353 evoked potent relaxation of isolated canine bladder strips. Furthermore, the nonselective β-AR antagonist bupranolol (Pietri-Rouxel and Strosberg, 1995) attenuated GW427353-evoked relaxations, demonstrating the involvement of β-AR stimulation in mediating these effects. Atenolol and ICI 118551, at concentrations shown to be effective in selectively blocking β1- and β2-ARs, respectively (Baker, 2005), were without effect. The present findings are in agreement with the study by Takeda et al. (2003) who showed potent relaxation of dog bladder strips with selective β3-, but not β1- or β2-AR agonists. Yamazaki et al. (1998) provided additional in vitro functional evidence using selective β-AR agonists and antagonists to show that β3-ARs mediate the bladder relaxation induced by β-AR agonists in the dog.
GW427353 also demonstrated the ability to relax canine detrusor in vivo as measured by a significant increase in the micturition volume threshold, i.e., an increase in bladder capacity, under conditions of acetic acid-evoked bladder irritation. No effects on voiding efficiency were observed, demonstrating that GW427353 did not detrimentally affect the ability of the bladder to void. Consistent with the in vitro findings in canine isolated detrusor strips, bupranolol blocked the GW427353-evoked increases in micturition volume threshold, suggesting the involvement of β-AR stimulation. However, the in vivo results must be interpreted with caution, because β1-AR- or β2-AR-selective antagonists were not tested in vivo, and GW427353 was not tested for selectivity at canine recombinant β-ARs; therefore, a contribution of β1-AR and or β2-AR activity cannot be ruled out. Based on plasma exposures of GW427353 after administration, the high plasma protein binding and low volume of distribution (data not shown) of this compound in the dog, drug levels may not have been sufficient at the 1 mg/kg dose, especially at the level of the bladder tissue, to evoke a similar pharmacological effect to 3 mg/kg i.v. on bladder function. However, GW427353 evoked effects on cardiovascular parameters, characterized by flushing, decreases in MAP, and a positive chronotropic effect, consistent with β3-AR stimulation in the dog, at both doses tested (Tavernier et al., 1992; Shen et al., 1996). Differences in the distribution of GW427353 to visceral organs and peripheral circulation or differences in β3-AR density and physiological function at these two sites may explain the differential magnitude of the effects of GW427353 on bladder versus the cardiovascular system. GW427353-evoked cardiovascular responses were only partially attenuated by bupranolol. It has been reported that complete sinoaortic denervation or combined ganglionic, cholinergic, and adrenergic (β1- and β2-AR) blockade is required to completely block β3-AR agonist-evoked increases in HR in the dog. This indicates that the β3-AR agonist effects on cardiovascular function in this species are complex, and that the effects on MAP and HR involve a variety of direct and indirect mechanisms (Tavernier et al., 1992; Shen et al., 1996; Donckier et al., 2001). In addition, the apparent β3-AR-mediated cardiovascular effects in dogs seem to be species-specific, because reports suggest selective β3-AR agonism in nonhuman primates and humans does not result in similar cardiovascular effects (Shen et al., 1996; Larsen et al., 2002).
SR59230A, which has been shown to have antagonistic properties at the β3-AR (Manara et al., 1996), was used in the present study in an attempt to provide further supporting evidence for the mechanism of action of GW427353. SR59230A evoked responses alone in the isolated tissue studies, suggesting β3-AR agonist activity. Partial β3-AR agonist activity of SR59230A has been reported previously in guinea pig gastrointestinal tissues (Horinouchi and Koike, 2001) and at the mouse β3-AR (Hutchinson et al., 2005). We also observed partial agonist activity of SR59230A on CHO cells expressing human recombinant β3-ARs (EC50 = 3.4 nM, intrinsic activity = 0.28; data not shown).
In summary, the present study has demonstrated the ability of a novel selective β3-AR agonist GW427353 to evoke bladder relaxation and to facilitate bladder storage mechanisms in the dog. Because β3-ARs are postulated to play an important role in the control of bladder smooth muscle tone in both dog and human, these results support the premise that β3-AR agonists such as GW427353 represent a useful clinical strategy for the treatment of overactive bladder.
Footnotes
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.107.125757.
-
ABBREVIATIONS: AR, adrenergic receptor; GW427353, (R)-3′-[[2-[[2-(3-chlorophenyl)-2-hydroxyethyl]amino]ethyl]amino]-[1,1′-biphenyl]-3-carboxylic acid; CHO, Chinese hamster ovary; ICYP, iodocyanopindolol; SR59230A, 1-(2-ethylphenoxy)-3-[[(1S)-1,2,3,4-tetrahydro-1-naphthalenyl]amino]-(2S)-2-propanol; DMSO, dimethyl sulfoxide; ICI 118551, (±)-1-[2,3-(dihydro-7-methyl-1H-inden-4-yl)oxy]-3-[(1-methylethyl)amino]-2-butanol; MAP, mean arterial blood pressure; HR, heart rate.
- Received May 15, 2007.
- Accepted July 11, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}