


Abstract
Cytochrome P450 enzymes belonging to the CYP2D subfamily have been shown to be one of determinants of the polymorphic drug oxidations in the human and the rat. Debrisoquine 4-hydroxylation is a typical reaction catalyzed by these enzymes. However, various strains of mice were observed to have much lower debrisoquine 4-hydroxylase activity than Wistar rats, whereas other monooxygenase activities in mice toward bunitrolol, propranolol, imipramine and amitriptyline, which are mediated by the CYP2D enzymes in the rat, were comparable to those of the rats. Immunoblot analysis of mouse liver microsomes with an antibody raised against a rat CYP2D enzyme indicated that the mouse liver contained a P450 enzyme(s) immunochemically related to the rat CYP2D enzyme. The antibody inhibited propranolol ring-hydroxylase and imipramine 2-hydroxylase activities, as well as testosterone 16α-hydroxylase activity, a typical reaction of mouse CYP2D9, but not debrisoquine 4-hydroxylase activity in mouse liver microsomes. We partially purified a P450 enzyme (designated P450 ML2d) from livers of male ddY mice by monitoring the cross-reactivity with the antibody. The partially purified enzyme was indicated to belong to the CYP2D subfamily from its N-terminal amino acid sequence, but the homology of the sequence to other CYP2D enzymes of the mouse (CYP2D9–11) was 62%, suggesting that P450 ML2d is a novel P450 enzyme. P450 ML2d had the oxidation activities for the rat CYP2D-substrates, such as propranolol 4-hydroxylation and imipramine 2-hydroxylation, in higher rates than those of the microsomes, but did not exhibit debrisoquine 4-hydroxylase activity. Our result is the first finding that a mouse CYP2D enzyme also metabolizes substrates for the rat CYP2D enzyme, in addition to steroids, but the enzyme had a limited specificity for the substrates of the CYP2D enzymes of the rat and the human.
The oxidative metabolism of various β-blockers and antidepressants exhibits debrisoquine-type genetic polymorphism in the human. A P450 enzyme, CYP2D6, has been shown to be a determinant of the polymorphic drug oxidations (Murray, 1992). The rat has an orthologous enzyme, CYP2D1, whose substrate specificity is similar to that of CYP2D6 (Nedelcheva and Gut, 1994). We have found that another P450 enzyme in the CYP2D subfamily, CYP2D2, had catalytic activities similar to CYP2D1 (Suzuki et al., 1992; Ohishi et al., 1993). Other four genes and/or proteins of the CYP2D subfamily (CYP2D3, 4, 5 and 18) were also found in rats (Nelson et al., 1996). In connection with the genetic polymorphism, the DA strain rat, which is functionally deficient in CYP2D1, was established as a poor metabolizer animal model of debrisoquine 4-hydroxylation (Al-Dabbagh et al., 1981;Kahn et al., 1985), although some discrepancy was recognized from the kinetic and the gene analysis with the strain (Matsunagaet al., 1989; Barham et al., 1994). However, knowledge on the metabolism of substrates for CYP2D6 and/or CYP2D1 in other rodents was limited. A study with liver microsomes from 12 mouse strains (Adams et al., 1991) indicated that the average formation ratio for two oxidative metabolites of metoprolol, a typical substrate of CYP2D6, was similar to that of human liver microsomes. Although five genes encoding the CYP2D enzymes have been isolated from mouse liver (Cyp2d9–13, Nelson et al., 1996), it has not been studied whether the CYP2D enzymes have ability to metabolize substrates for CYP2D6 and/or CYP2D1.
In our preliminary study, we observed that hepatic microsomal debrisoquine 4-hydroxylase activity was much lower in the mouse than in the rat, although the oxidation activities toward some β-blockers were similar. The purpose of our study is to clarify the cause of an apparent difference in the monooxygenase activity between the mouse and the rat. We thus investigated participation of the CYP2D enzymes in debrisoquine 4-hydroxylase and related monooxygenase activities in mouse liver microsomes.
Materials and Methods
Chemicals.
Debrisoquine and 4-hydroxydebrisoquine as hemisulfates were obtained from Hoffmann-La Roche (Basel, Switzerland); bunitrolol and 4-hydroxybunitrolol as hydrochlorides from Nippon Boehringer Ingelheim Co. (Hyogo, Japan); 4-hydroxypropranolol hydrochloride from the Sumitomo Chemical Co. (Osaka, Japan); 2-hydroxyimipramine from Geigy (Basel, Switzerland); E-10- and Z-10-hydroxyamitriptylines as hydrochlorides from H. Lundbeck A/S (Copenhagen, Denmark). Propranolol, imipramine and amitriptyline as hydrochlorides, and testosterone, 2α- and 16α-hydroxytestosterones were purchased from the Sigma Chemical Co. (St. Louis, MO). 5-Hydroxy- and 7-hydroxypropranolols were synthesized as hydrochlorides according to the method of Oatis et al.(1981). 6β- and 7α-hydroxytestosterones were purchased from Steraloids Inc. (Wilton, NH); Sepharose 4B from Pharmacia Fine Chemicals (Uppsala, Sweden); Emulgen 911 from Kao Atlas Co. (Tokyo, Japan); hydroxyapatite from Bio-Rad Laboratories (Richmond, CA); G-6-P, G-6-PDH and NADPH from the Oriental Yeast Co., Ltd. (Tokyo, Japan). All other chemicals and solvents used were of analytical grade.
Preparation of hepatic microsomes.
Male and female C57BL/6, DBA/2 and ddY mice (2 mo old) were obtained from the Japan SLC Inc. (Shizuoka, Japan). Male Wistar rats, Hartley guinea pigs and Japanese White rabbits (2 mo old) were obtained from Takasugi Experimental Animals (Saitama, Japan). The animals were housed in air-conditioned rooms (25°C) under a 12 hr light-dark cycle for 1 wk before use. Food (commercially available pellet, the Oriental Yeast Co., Ltd.) and water were given ad libitum. Liver microsomal fractions were prepared according to the method of Omura and Sato (1964). Protein concentrations were assayed by the method of Lowry et al.(1951).
Assay of microsomal enzymatic activities.
Debrisoquine 4-hydroxylase (Masubuchi et al., 1991), bunitrolol 4-hydroxylase (Suzuki et al., 1991), propranolol 4-, 5-, and 7-hydroxylase (Masubuchi et al., 1993), imipramine 2-hydroxylase (Chiba et al., 1988), amitriptylineE- and Z-10-hydroxylase (Fujita et al., 1989) and testosterone oxidase activities (Hayashi et al., 1986) of the microsomes were assayed by the HPLC methods previously reported. Substrate concentrations used were: debrisoquine, 2 mM; bunitrolol, 10 μM; propranolol, 1 mM; imipramine, 50 μM; amitriptyline, 500 μM; testosterone, 250 μM. In kinetic studies, substrate concentration ranges used were: debrisoquine, 1 to 2000 μM; bunitrolol, 0.5–1000 μM.
Immunochemical analysis with the antibody against the rat CYP2D enzyme.
A P450 enzyme belonging to the CYP2D subfamily (P450BTL) was purified from liver microsomes of male Sprague-Dawley rats (2 mo old, the Japan SLC. Inc.) by monitoring bunitrolol 4-hydroxylase activity (Suzuki et al., 1992). An N-terminal amino acid sequence of P450BTL (GLLIGXDLMAVVXFXAIXLL) was very similar to CYP2D2. P450BTL exhibited high debrisoquine 4-hydroxylase activity (2.22 and 5.63 nmol/min/nmol P450 at substrate concentrations of 50 μM and 2 mM, respectively). Thus, P450BTL was indicated to belong to the CYP2D subfamily. Polyclonal antibody against the enzyme was raised in female Japanese White rabbits (Takasugi Experimental Animals), and it suppressed debrisoquine 4-hydroxylase activity in rat liver microsomes almost completely. Immunoblot analysis revealed that the antibody cross-reacted with none of the purified rat liver P450 enzyme belonging CYP1A, 2B, 2C, 2E or 3A subfamily. Immunoblot analysis of liver microsomes obtained from various animal species was performed as previously reported (Laemmli, 1970; Guengerich et al., 1982). Briefly, microsomal proteins (5.0 μg) were separated by SDS-PAGE with a 10% polyacrylamide gel. The proteins on the gel were transferred to a nitrocellulose membrane (Bio-Rad Laboratories), followed by treatment of the membrane with the antibody against the CYP2D enzyme. In immunoinhibition studies, microsomes were preincubated with various amounts of the anti-CYP2D antibody or preimmune serum at 25°C for 30 min, followed by adding other components of the incubation mixture and assay of the oxidation activities.
Purification and characterization of a P450 enzyme in the CYP2D subfamily from mouse liver microsomes.
A P450 enzyme was purified from mouse liver by the previously reported methods used for the rat enzymes (Suzuki et al., 1992; Ohishi et al., 1993) with modifications. Cross-reactivities with the antibody directed to the rat CYP2D enzyme (P450BTL) described above were used as an index of the purification. Liver microsomes prepared from male ddY mice were solubilized with sodium cholate (3 mg/mg microsomal protein) and precipitated with polyethyleneglycol (7–16%, w/v). The precipitation was applied to an ω-aminooctyl-Sepharose 4B column (5 cm i.d. × 35 cm) equilibrated with 100 mM potassium phosphate buffer (pH 7.2) containing 20% (v/v) glycerol, 1 mM EDTA, 0.5 mM dithiothreitol and 0.5% (w/v) sodium cholate. The column was washed with the same buffer, and P450 enzymes, which were monitored with the absorbance at 417 nm, were eluted with the buffer containing 0.4% sodium cholate and 0.1% (w/v) Emulgen 911. This fraction was subjected to HPLC using a DEAE-5PW column (2.15 cm i.d. × 15 cm, Tosoh, Tokyo, Japan). P450 enzymes were eluted with 20 mM Tris-acetate buffer (pH 7.5) containing 20% glycerol and 0.4% Emulgen 911 at a flow rate of 1.0 ml/min, and followed by a linear gradient of sodium acetate from 0 to 1.0 M in this buffer over 40 min. Among three fractions with absorption at 417 nm obtained, a second fraction contained the protein that was recognized by the anti-CYP2D antibody. Because SDS-PAGE indicated that the fraction contained proteins which were not recognized by the antibody, it was further subjected to HPLC with a Protein pack G-SP column (8.2 mm i.d. × 75 mm, Waters Assoc., Milford, MA) equilibrated with 20 mM sodium phosphate buffer (pH 6.5) containing 20% glycerol and 0.4% Emulgen 911. P450 enzymes were eluted with a linear gradient of sodium acetate from 0 to 0.5 M in this buffer at a flow rate of 1.0 ml/min over 40 min. Most of the proteins were eluted as a pass-through fraction of the column, followed by elution of three fractions with a gradient of sodium acetate. The first fraction contained a single protein band in SDS-PAGE, which was recognized by the anti-CYP2D antibody. To remove Emulgen 911, it was applied to a hydroxyapatite open column (7.5 mm i.d. × 75 mm, Bio-Rad Laboratories) equilibrated with potassium phosphate buffer (pH 7.4) containing 20% glycerol and 0.2% sodium cholate. The column was washed with the same buffer until the absorption of Emulgen 911 at 280 nm in the eluate disappeared, and the P450 was eluted with 350 mM potassium phosphate buffer (pH 7.4) containing 20% glycerol and 0.2% sodium cholate. A final preparation thus obtained was designated tentatively P450 ML2d. The recovery of the P450 ML2d from microsomes was 0.04%, and the specific content was 5.04 nmol/mg protein. An apparent molecular mass of P450 ML2d was 50 kDa on SDS-PAGE.
Other methods.
P450 content was measured spectrally by the method of Omura and Sato (1964). ω-Aminooctyl-Sepharose 4B was prepared according to the method of Guengerich and Martin (1980). SDS-PAGE was performed by the method of Laemmli (1970) and the proteins on the slab gels were stained with alkaline AgNO3solution. An N-terminal amino acid sequence was analyzed according to the reported method (Matsudaira, 1987). NADPH-P450-reductase was purified from liver microsomes of male Sprague-Dawley rats by the method of Yasukochi and Masters (1976). Enzyme activities in a reconstituted system were determined according to the microsomal assay method, but the following components were added instead of the microsomal protein: P450 ML2d, 50 pmol; NADPH-P450-reductase, 0.5 U; dilauroylphosphatidylcholine, 5 μg; sodium cholate, 0.1 mg.
Data analysis.
Enzyme kinetic parameters (Km and V max) were analyzed according to a nonlinear least squares regression analysis based on a simplex method (Yamaoka et al., 1981). Best fittings of the data were performed by weighting them with the reciprocal of their square. Statistical significance was calculated by the Student’s t test.
Results
Selective deficiency of debrisoquine 4-hydroxylation in mice.
Debrisoquine 4-hydroxylase and bunitrolol 4-hydroxylase activities, both of which mainly reflect the functions of rat and human P450 isozymes belonging to the CYP2D subfamily (Suzuki et al., 1992), were determined with liver microsomes from two mouse inbred strains, C57BL/6 and DBA/2, and an outbred strain, ddY, of both sexes. Compared with the reported values of the rat (Suzuki et al., 1991), debrisoquine 4-hydroxylase activities in mice were much lower than those in Wistar rats for the corresponding sexes (4–7% in males, 14–16% in females, fig. 1A). The activities in the mouse were comparable to those in the DA rats, which is known as a poor metabolizer animal model for debrisoquine 4-hydroxylation (Al-Dabbagh et al., 1981; Kahn et al., 1985). No major strain difference was observed in mice, whereas the activities in C57BL/6 and DBA/2 mice revealed a small but significant sex difference (female > male). Bunitrolol 4-hydroxylase activities were also lower in mice than in Wistar rats (fig.1B). However, the extent of the differences was smaller in bunitrolol 4-hydroxylation than in debrisoquine 4-hydroxylation (22–49% in males, 46–105% in females), and the activity in female C57BL/6 or DBA/2 mouse was similar to that in female Wistar rat. The activities of the mouse were much higher than those of the DA rat.

Debrisoquine 4-hydroxylase and bunitrolol 4-hydroxylase activities in liver microsomes of rats and mice. Liver microsomes were obtained from Wistar rats (R/WIS), Dark Agouti rats (R/DA), C57BL/6 mice (M/C57), DBA/2 mice (M/DBA) and ddY mice (M/ddY) of males (░) and females (▨). Debrisoquine 4-hydroxylase (A) and bunitrolol 4-hydroxylase (B) activities were assayed at a substrate concentrations of 2 mM and 10 μM, respectively. Each value represents the mean ± SE of four or five determinations. Data of the rats were quoted from Suzuki et al. (1991). a,aaSignificantly different from males (P < .05, P < .01, respectively).
Debrisoquine 4-hydroxylation in mouse liver microsomes was analyzed kinetically as a single Michaelis-Menten equation (table1). Compared with the parameters obtained in the Wistar rat, the low-Km phase was deficient in the mouse. The Km values in the mouse were close to the value of high-Km phase in the rat, and theV max values were lower than the value of this phase in the rat. Bunitrolol 4-hydroxylation showed monophasic kinetics in the mouse (table 1) as well as in the rat. TheKm values in the mouse were 10-fold higher than the reported values in Wistar rats, and theV max values were close between the two animal species (Suzuki et al., 1991).
Kinetic parameters for debrisoquine 4-hydroxylase and bunitrolol 4-hydroxylase activities in liver microsomes of male mice and rats
Other monooxygenase activities mediated by rat CYP2D enzyme were compared between male Wistar rats and male C57BL/6 mice. Propranolol 4-, 5- and 7-hydroxylase activities (fig.2A) and amitriptylineE-10-hydroxylase activity (fig. 2B) in the mouse were significantly lower than those in the rat, and amitriptylineZ-10-hydroxylase activity in the mouse was higher than that in the rat. However the extents of the differences were within 50% of the corresponding activities. Imipramine 2-hydroxylase was not significantly different between the two animal species (fig. 2B).
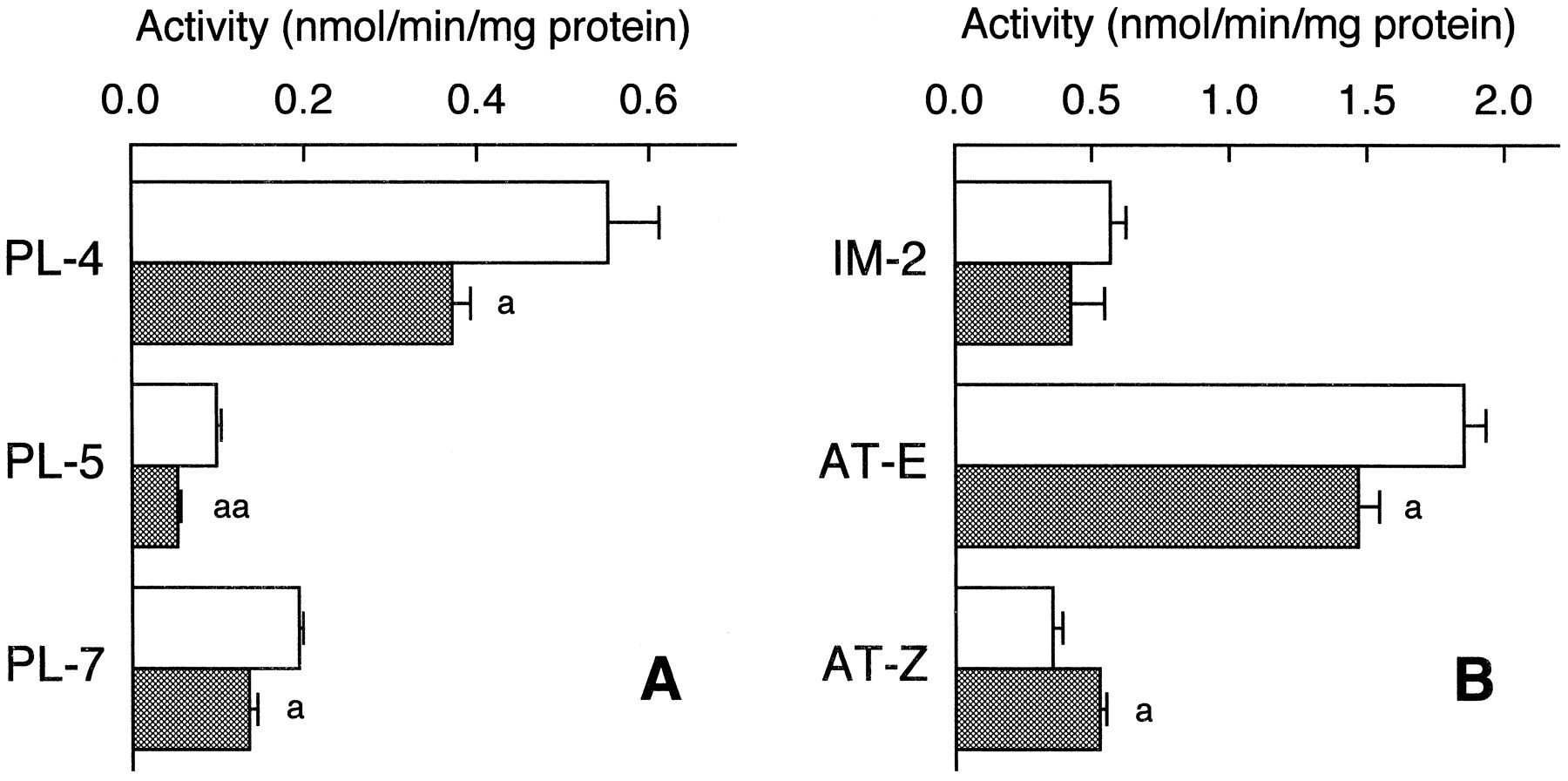
Oxidation activities toward propranolol, imipramine and amitriptyline in liver microsomes of rats and mice. Liver microsomes were obtained from male Wistar rats (□) and male C57BL/6 (▩) mice. Propranolol 4-, 5- and 7-hydroxylase (PL-4, PL-5 and PL-7, respectively), imipramine 2-hydroxylase (IM-2), and amitriptylineE-10- and Z-10-hydroxylase (AT-E and AT-Z, respectively) were assayed at substrate concentrations of 1 mM, 100 μM and 500 μM for propranolol, imipramine and amitriptyline, respectively. Each value represents the mean ± S.E. of three determinations. a,aa Significantly different from Wistar rats (P < .05, P < .01, respectively).
Immunoblot analysis of microsomes with the anti-CYP2D antibody.
Immunoblot analysis of liver microsomal fractions obtained from male C57BL/6, DBA/2 and ddY mouse, along with male Wistar rats, Japanese White rabbits and Hartley guinea pigs revealed that proteins which were immunoreactive with rat CYP2D enzyme exist in microsomes from all of the animal species examined (fig.3). This indicates that livers of the animal species other than the rat contain a P450 enzyme(s) immunochemically related to the rat CYP2D enzyme. The staining intensities were similar between the species except for the rabbit with a faint band, whereas the molecular masses of the bands were slightly different among the species.

Immunoblot analysis of liver microsomal proteins from various animal species with anti-rat CYP2D antibody. Liver microsomes (5 μg) from Wistar rats (A), Japanese White rabbits (B) and Hartley guinea pigs (C), C57BL/6 mice (D), DBA/2 mice (E) and ddY mice (F) of males were applied to the wells. The microsomes were electrophoresed on a 10% slab gel and transferred to a nitrocellulose membrane. The nitrocellulose was treated with the antibody against the rat CYP2D enzyme.
Immunoinhibition of mouse liver microsomal monooxygenase activities by the antibody directed to the CYP2D enzyme of rats.
The antibody against the CYP2D enzyme purified from the rat liver inhibited debrisoquine 4-hydroxylase and bunitrolol 4-hydroxylase activities in rat liver microsomes almost completely (Suzuki et al., 1992). Because a microsomal protein from mouse liver cross-reacted with the antibody in the immunoblot analysis, we attempted to immunoinhibit oxidation activities in the microsomes. The anti-CYP2D antibody did not inhibit debrisoquine 4-hydroxylase activity in liver microsomes of male C57BL/6 or ddY mouse (fig. 4A and B), whereas the antibody inhibited bunitrolol 4-hydroxylase activities in both strains of mouse (fig. 4C and D) in a concentration-dependent manner like in the rat (Suzuki et al., 1992). Similar immunoinhibition was observed in propranolol 4-, 5-, 7-hydroxylase, and imipramine 2-hydroxylase activities in liver microsomes from male C57BL/6 mice (fig. 5).

Effects of anti-rat CYP2D antibody on debrisoquine 4-hydroxylase and bunitrolol 4-hydroxylase activities in mouse liver microsomes. Liver microsomes obtained from male C57BL/6 mice (A, C) and male ddY mice (B, D) were preincubated with the antibody against the rat CYP2D enzyme (•) or preimmune serum (○), followed by assay of debrisoquine 4-hydroxylase (A, B) and bunitrolol 4-hydroxylase (C, D) activities. Results are expressed as per cent of the control activities obtained without the antibody. Each value represents the mean of two determinations.

Effects of anti-rat CYP2D antibody on oxidation activities toward propranolol and imipramine in mouse liver microsomes. Liver microsomes obtained from male C57BL/6 mice were preincubated with the antibody against the rat CYP2D enzyme, followed by assay of propranolol 4- (•), 5- (○), 7- (▴) hydroxylase and imipramine 2-hydroxylase (▵) activities. Results are expressed as per cent of the control activities obtained without the antibody. Each value represents the mean of two determinations.
Catalytic activities of a partially purified CYP2D enzyme, P450 ML2d.
Figures 6 and 7 show SDS-PAGE and the N-terminal amino acid sequence of P450 ML2d along with the sequences of CYP2D proteins reported previously, respectively. The sequence of P450 ML2d was different from that of any P450 listed, and showed a homology of 62% with all enzymes, indicating that P450 ML2d belongs to the CYP2D subfamily. P450 ML2d did not have debrisoquine 4-hydroxylase activity in the reconstituted system (table2). It had propranolol 4- and 5-hydroxylase activities higher than the corresponding activities in the microsomes, whereas its 7-hydroxylase activity could not be detected. P450 ML2d also exhibited high imipramine 2-hydroxylase activity in this system.

SDS-PAGE of partially purified P450 ML2d. P450 ML2d (5 pmol) was electrophoresed on a 10% slab gel. Std represents molecular mass standards including phosphorylase b (94 kDa), bovine serum albumin (67 kDa), ovalbumin (43 kDa) and carbonic anhydrase (30 kDa). The proteins on the gel were stained with alkaline AgNO3 solution.

Comparison of N-terminal amino acid sequence of P450 ML2d with those of the mouse CYP2D enzymes. The sequences for CYP2D9, CYP2D10 and CYP2D11 were quoted form Wong et al.(1987), Ichikawa et al. (1989), Wong et al.(1989), respectively, all of which were deduced from their complementary DNAs. X shows an unidentified amino acid residue.
Catalytic activities of P450ML2d in the reconstituted system
Participation of the mouse CYP2D enzyme in testosterone oxidation.
It has been reported that a mouse P450 enzyme in the CYP2D subfamily, CYP2D9, is involved in testosterone 16α-hydroxylation (Wong et al., 1989). P450 ML2d showed not only testosterone 16α-hydroxylase but also 6β- and 7α-hydroxylase activities in the reconstituted system (table 2). The antibody against the rat CYP2D enzyme inhibited testosterone 16α-hydroxylase activity in a concentration-dependent manner, but not the 6β- or 7α-hydroxylase activity in liver microsomes of male ddY mice (fig. 8).

Effects of anti-rat CYP2D antibody on testosterone hydroxylase activities in mouse liver microsomes. Liver microsomes obtained from male C57BL/6 mice were preincubated with the antibody against the rat CYP2D enzyme, followed by assay of testosterone 6β- (•), 7α- (○) and 16α-hydroxylase (▴) activities. Results are expressed as per cent of the control activities obtained without the antibody. Each value represents the mean of two determinations.
Discussion
Debrisoquine 4-hydroxylase activities in liver microsomes from three strains of mice were much lower than the values in Wistar rats. The lower activity could be explained kinetically as deficiency of a low-Km phase and a lowerV max value of a high-Km phase in the mouse. However, other monooxygenase activities in the mouse toward β-blockers and antidepressants as substrates, which have been shown to be mediated by the CYP2D in the rat (Suzuki et al., 1992; Masubuchiet al., 1993; 1995), were comparable to those in the rat. Thus, we tested effects of the antibody against the rat CYP2D enzyme on the oxidation activities in the mouse, because the antibody recognized proteins corresponding to P450 in mouse liver microsomes in the immunostaining analysis. The antibody inhibited the oxidation activities toward bunitrolol, propranolol and imipramine, but not debrisoquine 4-hydroxylation. These observations suggested that a mouse P450 enzyme(s) immunochemically related to the rat CYP2D enzyme did not mediate debrisoquine 4-hydroxylation, resulting in apparent selective lower activity in this reaction. Thus, at least following two possibilities are considered as the cause of the lower debrisoquine 4-hydroxylase activity in the mouse: 1) none of the mouse CYP2D enzymes catalyzes the reaction, and another P450 enzyme(s) that belongs to other family and/or subfamily than CYP2D mediates the reaction with a very low activity; 2) a mouse CYP2D enzyme(s) which is not recognized by the antibody has debrisoquine 4-hydroxylase activity but the activity is very low.
We conducted the purification of a P450 enzyme from mouse livers by tracing the cross-reactivity with the antibody directed to the rat CYP2D enzyme, yielding a partially purified P450 enzyme in the CYP2D subfamily (termed P450 ML2d) judging from its N-terminal amino acid sequence. P450 ML2d did not have debrisoquine 4-hydroxylase, but the other oxidation activities mediated by the CYP2D enzyme in the rat, such as propranolol 4-hydroxylation and imipramine 2-hydroxylation, were observed in higher rates than the corresponding values of the microsomes. These results mean that propranolol but not debrisoquine is a substrate for the partially purified CYP2D enzyme.
Five genes encoding the CYP2D enzymes (Cyp2d9–13) have been isolated from the mouse (Nelson et al., 1996) and expression of two proteins (CYP2D9 and 10) in the liver has been confirmed (Wonget al., 1987; 1989; Ichikawa et al., 1989;Sueyoshi et al., 1995). The former is expressed only in male mice and is involved in testosterone 16α-hydroxylation, whereas the latter is expressed in both sexes, and is not involved in the reaction. Judging from the immunochemical cross-reactivity and the homology of N-terminal amino acid sequence, P450 ML2d was thought to be a novel P450 enzyme belonging to CYP2D subfamily. The antibody against the rat CYP2D enzyme inhibited testosterone 16α-hydroxylase activity in mouse liver microsomes, suggesting that the antibody recognizes CYP2D9 and inhibits its oxidation activity, although P450 ML2d may also be involved in the reaction because the activity was obtained in the reconstituted system. It has not been known whether CYP2D9 and/or 2D10 are involved in debrisoquine 4-hydroxylase and related monooxygenase activities that are mediated by the rat CYP2D enzymes. In the present study, we could not exclude a possibility that CYP2D9 and/or CYP2D10 have debrisoquine 4-hydroxylase activity, because cross-reactivities of the antibody used in the present study with these enzymes have not been directly tested.
There are many examples that different P450 species (subfamily) are responsible for an oxidative reaction in different animal species (Nedelcheva and Gut, 1994), i.e., structurally and immunochemically related P450 enzymes in different animal species have different substrate specificities. These species differences in substrate specificities of the P450 enzymes make difficult to extrapolate the data of the drug metabolism from experimental animals to humans. In summary, the present result is the first finding that a mouse CYP2D enzyme(s) also metabolizes substrates for the rat CYP2D enzyme, in addition to steroids. Because the CYP2D enzyme(s) of the mouse was shown to have a limited substrate specificity as compared to those of the rat, the rat is considered to be a better experimental animal species than the mouse for a preliminary screening of the CYP2D-dependent- and polymorphic human drug metabolism. Debrisoquine 4-hydroxylase has been used as an index for the CYP2D enzyme, but is not appropriate for all animal species. Further analysis of mouse CYP2D enzymes may provide useful suggestion on determinants of substrate specificity of the CYP2D subfamily.
Footnotes
-
Send reprint requests to: Dr. Shizuo Narimatsu, Associate Professor, Laboratory of Biopharmaceutics, Faculty of Pharmaceutical Sciences, Chiba University, 1–33 Yayoi-cho, Inage-ku, Chiba 263, Japan.
- Abbreviations:
- P450 or CYP
- cytochrome P450
- DA
- Dark Agouti
- G-6-P
- glucose 6-phosphate
- G-6-PDH
- glucose 6-phosphate dehydrogenase
- SDS-PAGE
- sodium dodecyl sulfate-polyacrylamide gel electrophoresis
- Received December 9, 1996.
- Accepted May 21, 1997.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}