

Abstract
The cloning and expression of genes encoding for the human neuronal nicotinic acetylcholine receptors (nAChRs) has opened new possibilities for investigating their physiological and pharmacological properties. Cells (HEK 293) stably transfected with two of the major brain subunits, α4 and β2, were characterized electrophysiologically using the patch-clamp technique. Fast application of the natural ligand ACh can evoke currents up to 3500 pA, with an apparent affinity (EC50) of 3 μm and a Hill coefficient of 1.2. The rank order of potency of four nAChR ligands to activate human α4β2 receptors is (−)-nicotine > ACh > (−)-cytisine > ABT-418. At saturating concentrations, the efficacy of these ligands is ABT-418 ≫ (−)-nicotine > ACh ≫ (−)-cytisine > GTS-21 (previously named DMXB). Coapplication of 1 μm ACh with known nAChR inhibitors such as dihydro-β-erythroidine and methyllycaconitine reversibly reduces the current evoked by the agonist with respective IC50 values of 80 nm and 1.5 μm. The current–voltage relationship of human α4β2 displays a strong rectification at positive potentials. Experiments of ionic substitutions suggest that human α4β2 nAChRs are permeable to sodium and potassium ions. In the “outside-out” configuration, ACh evokes unitary currents (main conductance 46 pS) characterized by a very fast rundown. Potentiation of the ACh-evoked currents is observed when the extracellular calcium concentration is increased from 0.2 to 2 mm. In contrast, however, a reduction of the evoked currents is observed when calcium concentration is elevated above 2 mm.
Neuronal nicotinic acetylcholine receptors (nAChRs) constitute a family of cationic ligand-gated channels (Bertrand and Changeux, 1995; Galzi and Changeux, 1995) closely related to but distinct from the muscle nAChR. To date, eight neuronal α (α2–α9) and three neuronal β (β2–β4) subunits have been cloned in chick and rodents (Lindstrom, 1995; McGehee and Role, 1995) and also identified in humans (Anand and Lindstrom, 1990;Doucette-Stamm et al., 1993; Peng et al., 1994; Monteggia et al., 1995; for review, see Lindstrom, 1996). Furthermore, it was even shown that some neuronal subunits are transiently expressed in chick striated muscles during embryonic development (Corriveau et al., 1995)
The α4 and β2 subunits represent the most abundant nAChR subunits in chick and rat brains (Whiting et al., 1987; Schoepfer et al., 1988;Wada et al., 1989; Flores et al., 1992), and α4 mRNA is widely detected throughout the human cerebral cortex (Wevers et al., 1994). A good correlation exists between the high-affinity (−)-nicotine binding sites and the rodent brain regions expressing the α4 and β2 subunits (Clarke et al., 1985; Deutch et al., 1987; Swanson et al., 1987; Marks et al., 1992). Moreover, β2-containing nAChRs constitute the high-affinity binding sites for (−)-nicotine, as demonstrated in the β2 gene knock-out experiment in which (−)-nicotine binding to brain slices is fully abolished in transgenic mice (Picciotto et al., 1995). Thus, characterization of the physiological and pharmacological properties of the human α4β2 nAChR may be pertinent to a more complete understanding of the physiological effect of (−)-nicotine in human brain (Kellar et al., 1989; Marks et al., 1992; Sanderson et al., 1993; Lukas, 1995).
Alterations in the level/activity of brain nAChRs have been implicated in different neuropathologies such as Tourette’s syndrome (Silver and Sandberg, 1993), Parkinson’s disease (Baron, 1986; Janson et al., 1994), and even schizophrenia (Goff et al., 1992). In the brain of Alzheimer’s patients, the nAChR density is markedly reduced compared with that of age-matched controls (Whitehouse et al., 1986; Aubert et al., 1992). Moreover, the high-affinity binding of (−)-nicotine to brain regions that are believed to express mainly α4β2 nAChRs is extensively reduced in Alzheimer’s patients (Perry et al., 1995). The attention deficits observed in those patients have been attenuated by subcutaneous injection of (−)-nicotine (Jones et al., 1992), suggesting that (−)-nicotine, or other nAChR ligands (Decker et al., 1994a; Arendash et al., 1995; Arneric et al., 1995), could be used for the treatment of Alzheimer’s disease. In addition, several studies have demonstrated beneficial effects of (−)-nicotine on cognitive functions (Levin, 1992; Warburton, 1992; Picciotto et al., 1995).
Recently, a missense mutation in the human α4 neuronal nAChR subunit has been associated with the autosomal dominant nocturnal frontal lobe epilepsy (Steinlein et al., 1995). Other genetic investigations have linked the α4 nAChR subunit with another form of epilepsy: the benign familial neonatal convulsions (Beck et al., 1994; Schubert et al., 1994).
The human α4 and β2 cDNAs have recently been cloned (Monteggia et al., 1995) and stably transfected into HEK 293 cells (K177 clone;Sullivan et al., 1995; Gopalakrishnan et al., 1996). Herein we describe the electrophysiological properties of the human α4β2 nAChR using the whole-cell and single-channel patch-clamp techniques (Hamill et al., 1981).
MATERIALS AND METHODS
Cell transfection and culture. The cDNAs encoding the human α4 and β2 subunits were cloned into the BstXI site of the pRcCMV vector (Invitrogen, San Diego, CA) containing the genes conferring hygromycin or neomycin resistance, respectively (Gopalakrishnan et al., 1996). After transfection by lipofectamine (Gopalakrishnan et al., 1995), HEK 293 cells (K177 clone) were grown in DMEM with 10% fetal bovine serum plus the mixed antibiotic antimycotic at 37°C in an atmosphere of 95% air/5% CO2 at saturating humidity. Cells were cultured continuously in the presence of geneticin (250 μg/ml) and hygromycin (100 μg/ml) to avoid outgrowth of cells that do not express genes conferring the corresponding resistances and thereby also the α4 and β2 subunits. All cell culture products were from Life Technologies (Basel, Switzerland). Three to six days before electrophysiological recordings, the cells were incubated with trypsin for 3 min, mechanically dissociated, and seeded in 35-mm-diameter petri dishes (Nunc, Basel, Switzerland) at a density of ∼300 cells/dish. The same batch of K177 cells was cultured during a 15 month period without resampling from the frozen stock. No significant variation in the percentage of responsive cells or maximum ACh-evoked currents was observed during this time period. This suggests that the level of nAChR expression remains unchanged even after many cell cycles.
Electrophysiological recordings. The experiments were performed at room temperature. The standard bath solution for whole-cell recordings had the following composition (in mm): 120 NaCl, 5 KCl, 2 MgCl2, 2 CaCl2, 25 glucose, 10 Hepes; and 1 μmatropine (for blocking endogenous muscarinic receptors), pH 7.4 with NaOH. Borosilicate electrodes (2–8 MΩ) used for both whole-cell and “outside-out” recordings were filled with (in mm): 120 KF, 10 KCl, 5 NaCl, 2 MgCl2, 20 BAPTA, 10 Hepes, pH 7.4 with KOH. For ionic substitution experiments in the whole-cell configuration, MgCl2 was removed from the extracellular solution. The other modifications of the bath composition are given in the figure legends. The intracellular solution was modified as followed: NaCl and MgCl2 were omitted. Outside-out recordings were performed with an extracellular solution containing no MgCl2 and with Sylgard-coated electrodes to minimize the capacitance of the electrodes; the pipette was filled with the standard intracellular solution containing 2 mm MgCl2. Currents, recorded on isolated cells using an Axopatch 200A amplifier (Axon Instruments, Foster City, CA), were filtered on line at 1–2 kHz, digitized at 2–5 kHz, and stored on a personal computer equipped with an analog-to-digital converter (ATMIO-16D, National Instrument, Austin, Texas) and the DATAC package (Bertrand and Bader, 1986). Data were analyzed on a Macintosh Performa 5200 using the MacDATAC program. Fast superfusion of the cells was performed with a custom-made multibarrel (eight tubings) puffer, which allows drug application in the millisecond range (Puchacz et al., 1994; Gopalakrishnan et al., 1995;Bertrand et al., 1997), and fast exchange between the different solutions can be evaluated by ionic substitution during a steady response to a low ACh concentration (Fig. 1). All chemicals were from Sigma or Fluka (Buchs, Switzerland). Unless specified, the holding potential was −100 mV. All values are given as mean ± SEM.

Fast drug application using a multibarrel puffer. The time course for a complete solution exchange on a cell was determined by performing a sodium jump during a steady application of a low ACh concentration. The first “control” current (bold trace) was recorded in a standard medium containing 130 mm NaCl. A second current (light trace) was elicited by applying ACh in a solution containing 65 mmNaCl + 130 mm mannitol. Then the solution was switched for the standard medium (rectangle with diagonal lines), and the current was returned to the level of the control trace. The inset (bottom) presents with an expanded time scale the current rise time induced by the sodium jump. Complete solution exchange is performed within 20 msec.
RESULTS
Pharmacological properties of the human α4β2 nAChR
ACh, the endogenous ligand for human nAChRs, elicited currents in transfected cells at concentrations ≥0.1 μm(Fig. 2A). Maximal current recorded was up to −3500 pA, whereas the mean current amplitude of 20 cells challenged with 1 μm ACh was −824 ± 112 pA (holding potential, −100 mV), demonstrating the very substantial expression of α4 and β2 subunits in this cell line. Of 312 impaled cells, only 1.6% (five cells) did not present detectable currents. Data collected over a broad range of ACh concentrations (0.03–300 μm) were normalized and fitted with the empirical Hill equation, allowing the determination of an apparent half-effective concentration (EC50) of 3 μm and a Hill coefficient of 1.2 (Fig. 2B). Identical protocols were performed for three other agonists: (−)-nicotine, (−)-cytisine, and the recently characterized nAChR ligand ABT-418 (Arneric et al., 1994; Decker et al., 1994b). (−)-Nicotine was the most potent of the ligands investigated, with an EC50 of 1.6 μm(Fig. 2B), whereas (−)-cytisine and ABT-418 (with EC50 values of 11.6 and 13.9 μm, respectively) were less potent than either (−)-nicotine or ACh (Fig.3A). The efficacy of these agonists, and for the anabaseine-derived compound GTS-21 (previously named DMXB; Hunter et al., 1994) to activate α4β2 nAChRs, was determined at a saturating concentration (100 μm each but 300 μm for ABT-418; Fig. 3B–D). ABT-418 was the most efficacious of the agonists, eliciting 190 ± 15% of the ACh-evoked current (Fig. 3B,C). (−)-Nicotine was slightly more efficacious than ACh, but (−)-cytisine and GTS-21 behaved as very weak agonists at this nAChR (Fig. 3B,D). To further characterize the pharmacological profile of human α4β2 nAChRs, the effects of two well known competitive inhibitors of nAChRs, dihydro-β-erythroidine (DHβE) (Mulle and Changeux, 1990; Whiting et al., 1991) and methyllycaconitine (MLA) (Wonnacot et al., 1993), were investigated. MLA is a potent antagonist (in the nanomolar range) of the α7 homomeric nAChRs (Gopalakrishnan et al., 1995; Palma et al., 1996). Less than micromolar concentrations of DHβE completely blocked the ACh-evoked currents. Complete reversibility of inhibition was obtained within 10 sec (three cells; data not shown). The following inhibition protocol was then performed for DHβE: every 10 sec a 1 μm ACh-pulse was delivered for 1200 msec, and a DHβE jump was applied during the last 800 msec (Fig. 4,inset). The percentage of DHβE-induced inhibition has been plotted as a function of the logarithm of the antagonist concentrations, and the data were fitted with an empirical Hill equation (Fig. 4). The apparent half-inhibitory concentration (IC50) for DHβE was 80 nm.

Sensitivity of human α4β2 nAChRs toward ACh and (−)-nicotine. A, Currents evoked by increasing concentrations of ACh (pulse of 500 msec) in a cell held at −100 mV.B, Concentration–response curves for (−)-nicotine and ACh. Dose–response relationships were fitted with the empirical Hill equation y = 1/(1 + ((EC50/[agonist])n)). Half-effective concentrations (EC50) were then determined: 1.6 μm (n = 1.3) for (−)-nicotine (5 cells) and 3 μm (n = 1.2) for ACh (19 cells).

Agonists are characterized by different affinities and efficacies for human α4β2 nAChRs. A, Transfected HEK 293 cells were challenged with six to seven concentrations of each agonist (ranging from 0.3 to 300 μm). Data were then normalized against the largest evoked current and plotted on a semi-logarithm scale. Mean values were fitted with the empirical Hill equation (see Fig. 2 legend) giving EC50 values (μm) and Hill coefficient (n): (−)-cytisine (11.6, 1.5; 7 cells) and ABT-418 (13.9, 1.3; 13 cells).B, The efficacy of the agonist was determined at saturating concentration (100 μm each but 300 μm for ABT-418), with a holding potential of −100 mV. For accurate determination, the following protocol of sequential drug application (500 msec each, every 4.5 sec) was used: alternate applications of the agonists to be tested and ACh. A first set of measures was performed with ACh, (−)-nicotine, and ABT-418 (7 cells) and another one with ACh, (−)-cytisine and GTS-21 (7 cells). Currents were normalized as in A. ABT-418 evoked 190 ± 15% of saturating ACh current, whereas (−)-nicotine produced 112 ± 7%. In contrast, (−)-cytisine and GTS-21 can elicit only 16 ± 2% and 6 ± 2% of the ACh-evoked current, respectively. A typical example of the currents evoked on one cell by 100 μm ACh, (−)-nicotine and 100 μm ABT-418 is presented in C. A very weak current evoked by 100 μm GTS-21 is compared with the 100 μm ACh current for another cell in D.

Dihydro-β-erythroidine (DHβE) antagonizes the effect of ACh on the human α4β2 nAChRs. Every 10 sec, a 1200 msec pulse was delivered alternately with 1 μm ACh alone or with 1 μm ACh and increasing concentrations of DHβE (0–600 nm) in co-application for the last 800 msec (see an example of the recorded currents on one cell in the inset). The full block of the ACh-evoked current by 600 nm DHβE was totally reversed within 10 sec (not shown; 3 cells). The inhibitory effect of DHβE was measured at the end of the pulse of co-application (steady-state current) and normalized toward the value of the plateau amplitude of the preceding ACh-evoked current. Values were plotted against the concentrations of DHβE (on a logarithm scale) and fitted with the empirical Hill equation: y = 1/(1 + (([DHβE]/IC50)n)), where IC50 and n represent the half-inhibitory concentration and the Hill coefficient, respectively. The calculated IC50 value is 80 nm with an nvalue of 1.1 (n = 6).
Using the same methodological approach, MLA inhibited the functional activity of human α4β2 nAChRs with an apparent IC50 of 1.5 μm (Fig. 5), which is not significantly different from the value previously measured for the chick α4β2 nAChR (Drasdo et al., 1992). No detectable block of human α4β2 nAChRs was observed for concentrations of MLA below 0.5 μm. By analogy to observations made on other neuronal nAChRs (Palma et al., 1996; Harvey and Luetje, 1996), it is supposed that both DHβE and MLA act on the human α4β2 as competitive inhibitors. Confirmation of this mode of action, however, has not been examined further.

Methyllycaconitine (MLA) inhibits the ACh-evoked current at the human α4β2 nAChR in the micromolar range. A protocol identical to that used in Figure 4 was applied for the determination of the antagonistic properties of MLA. Traces corresponding to a 1 μm ACh-evoked current before and after a 6 μm MLA jump are presented in A. Normalized values were plotted as a function of the concentration of MLA represented on a logarithm scale (B) and fitted with the empirical Hill equation (see Fig. 4 legend). An IC50 of 1.5 μm (n = 1.8) was calculated from the mean ± SEM values collected on four cells.
As previously reported for chick (Bertrand et al., 1990) and for rat (Charnet et al., 1992) α4β2 nAChRs, the open-channel blocker hexamethonium (at concentrations higher than 10 μm) strongly inhibited the ACh-evoked current when co-applied with ACh (n = 8; data not shown).
Ionic permeability and voltage dependence of the human α4β2 nAChR
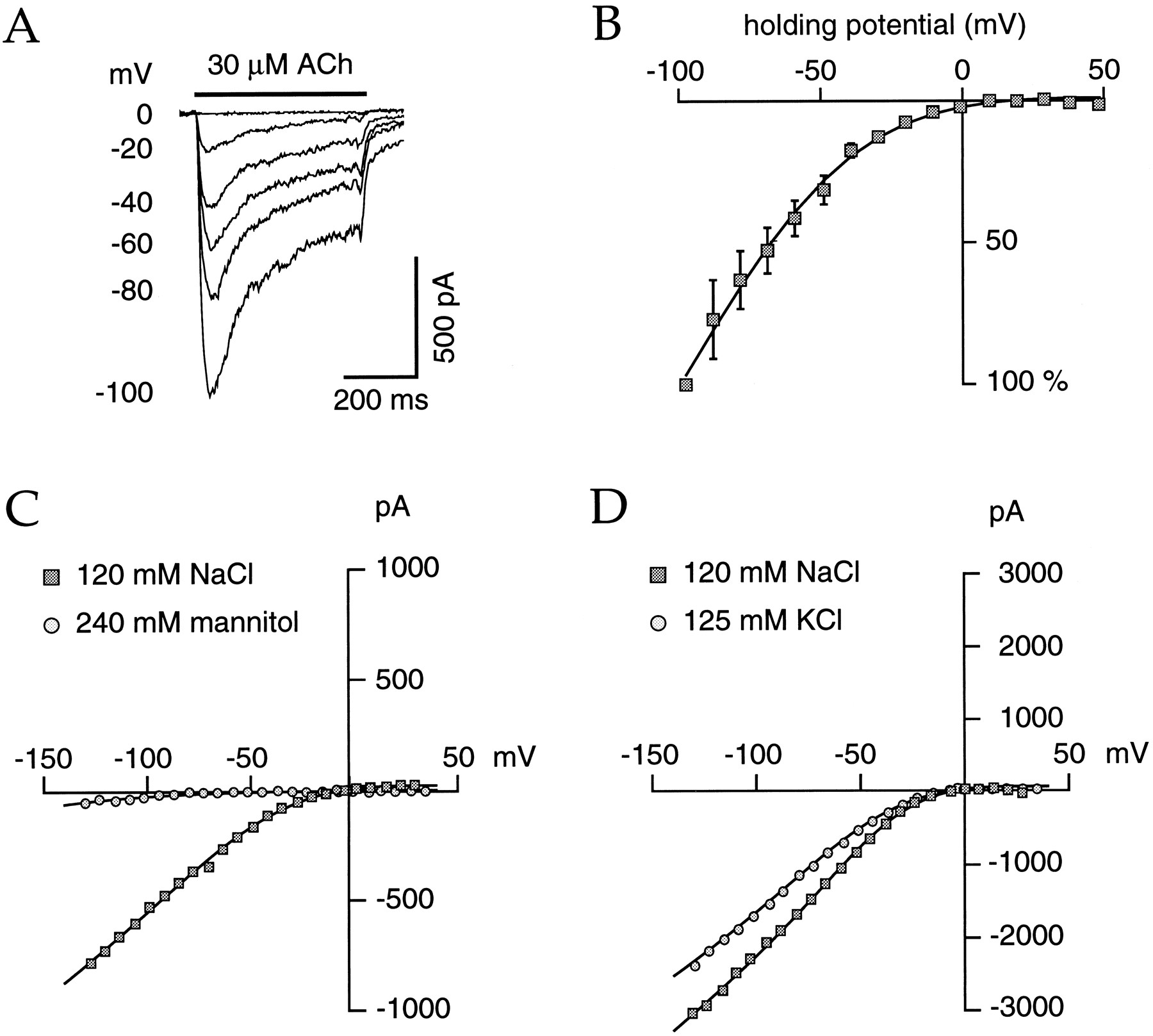
Determination of the human α4β2 nAChR current–voltage relationship was accomplished either with voltage step or voltage ramp protocols. Recordings of the ACh-evoked currents obtained over a broad voltage range are illustrated in Figure 6. In every cell tested, ACh failed to evoke detectable current for potentials above 0 mV. Figure 6B illustrates the mean amplitude currents obtained from three cells.

Human α4β2 nAChRs demonstrate strong rectification at positive potentials and are highly permeable to Na+ and K+ ions. ACh-evoked currents were recorded for holding potentials ranging from −100 to 40 mV with 10 mV steps every 4.5 sec in the standard extracellular solution; ACh was delivered 400 msec after the new set of the voltage. Apresents typical currents; for clarity, traces were omitted for all odd values. B, Currents were normalized against values measured at −100 mV. Mean values of three cells were plotted as a function of the holding potentials. Data were fit (solid line) by the following equation: y = (V − Er)/(1 + exp(α × (V − V1/2))), whereV is the holding voltage, Eris the current reversal potential, V1/2 is the potential for the half-current amplitude, and α is the slope factor. This formula corresponds to the product of the driving force and a Boltzmann equation. Optimal fit was obtained withV1/2 = −83.1 mV and α = 0.043. For the determination of the selectivity against cations, experiments using voltage ramps were performed with different extracellular solutions (C, D). The membrane was held at −10 mV, and a first ramp (from 40 to −140 mV in 400 msec) was applied in saline medium (without ACh) for the determination of the leak current. Four seconds later, 1 μm ACh was delivered for 800 msec, and the voltage ramp was applied again 200 msec after the beginning of ACh delivery. The leak current has been subtracted to the ACh-evoked current for all the data presented. For clarity, experimental current values are plotted every 20 points, corresponding to approximately one measurement every 7.5 mV. When extracellular Na+ chloride (NaCl) was replaced by mannitol, the quantity of ACh-evoked current was very weak (C), indicating a high permeability of the human α4β2 nAChR for Na+ ions (but see text). This nAChR is also highly permeable to potassium ions. When extracellular Na+ chloride is replaced by an equimolar concentration of potassium chloride, the amplitude of the ACh-evoked current is still large (D); note that the reversal potential for potassium ions switched from approximately −80 mV to 0 mV during this ionic substitution.
The strong rectification of these channels and the absence of significant outward currents at positive potentials prevented any reliable measurements of the ACh-evoked current reversal potential. This precluded clear cut determination of the ionic selectivity of this nAChR. Because intracellular Mg2+ has been shown to participate in the rectification mechanisms in different nAChR preparations (Mathie et al., 1990; Ifune and Steinbach, 1992; Sands and Barish, 1992; Albuquerque et al., 1995; Forster and Bertrand, 1995;Bonfante-Cabarcas et al., 1996), experiments were designed to reduce the intracellular free Mg2+ concentration; however, rectification was not modified by either removal of Mg2+from the intracellular medium or addition into the pipette of 5 mm CDTA, a specific chelator agent that was successfully used to overcome rectification of the homomeric α7 nAChR (Forster and Bertrand, 1995).
Substitution of the extracellular sodium by mannitol (Bertrand et al., 1993) almost completely abolished the ACh-evoked current (Fig.6C), whereas replacement of the extracellular sodium chloride by potassium chloride (Fig. 6D) or by sodium methanesulfonate (data not shown) induced no significant reduction of the ACh-evoked current. Taken together, these data indicate that as with other known nAChRs, these channels should be permeable to cations and that under physiological conditions, sodium ions are likely the major carrier ions flowing through the open nAChR channels.
Single-channel recordings
All outside-out patches were pulled in a magnesium-free solution, whereas pipette medium contained 2 mm MgCl2. Single-channel activities were regularly observed on withdrawal of the electrode from the whole-cell configuration (n = 75). This activity was observed in response to ACh and readily blocked by co-application of DHβE (600 nm; n = 3), thereby demonstrating that such elementary currents corresponded to multiple openings of human α4β2 nAChRs (Fig.7A). In all patches recorded, a predominant current amplitude was always observable at different holding potentials (Fig. 7B,D). Plots of the main single current amplitude as a function of the holding voltage (Fig. 7C,D) could be fitted with a straight line yielding a conductance of 46 pS and a reversal potential of ∼−7 mV. Single-channel properties of neuronal α4β2 nAChRs from chick and/or rat have been investigated previously (Ballivet et al., 1988; Papke et al., 1989; Cooper et al., 1991;Charnet et al., 1992). Only one conductance state of 20 pS has been described for the chick α4β2 nAChR (Ballivet et al., 1988; Cooper et al., 1991), whereas two or three (12, 22, and 34 pS) conductances were observed for the rat α4β2 nAChR (Papke et al., 1989; Charnet et al., 1992). Single-channel recordings on M10 cells expressing the chick α4β2 nAChR also revealed two conductance states (18 and 30 pS; Pereira et al., 1994). Recordings of the single-channel properties of the human α4β2 showed a predominant conductance of ∼46 pS. Occasionally, however, multiple levels were observed in some recordings, suggesting that channels can either undergo a rapid flickering or might display multiple conductances. Importantly, a rundown comparable to that reported originally for the chick and rat α4β2 nAChR (Ballivet et al., 1988; Charnet et al., 1992; Pereira et al., 1994) was observed in every patch analyzed (see below). Obviously, however, given the short time of recordings, we cannot eliminate the possibility of other conducting states occurring at very low frequency (Papke et al., 1989; Charnet et al., 1992). Many investigations have indicated that second transmembrane segments (TM-2) of the neuronal nAChRs subunits form the channel wall and determine the receptor ionic selectivity (for review, see Bertrand et al., 1993;Karlin and Akabas, 1995). Alignment of the TM-2 amino acid sequences of chick, rat, and human reveals an excellent conservation of this segment among the α4 and β2 subunits (Le Novère and Changeux, 1995;Lindstrom, 1996). Thus, differences in the main single-channel conductances observed between chick, rat, and human α4β2 nAChRs are not readily attributable to differences of the TM-2 segments. In addition, it is unlikely that differences in single-channel conductances result from the expression method of these subunits with either mammalian cell lines or Xenopus oocytes. Differences in channel conductance, however, could result from recording conditions, and namely the ionic strength used.

In the outside-out configuration, ACh-evoked single channels are blocked by DHβE, display a main conductance of 46 pS, and do not rectify at positive holding voltages. Intracellular solution (see Materials and Methods) contained 2 mmMgCl2. A high concentration of DHβE (600 nm; IC50 = 80 nm) completely inhibited the ACh-evoked single channels (A), demonstrating that these single-channel currents resulted from nAChR activation by ACh (n = 3). An outside-out patch stimulated by ACh at different holding potential clearly showed the voltage dependence of the ACh-evoked current and the predominant conductance observed in many other records (n = 46) (B). This patch contained at least two channels, but their activity disappeared before we were able to record at positive potentials. The closed (c) and opened (o) states are indicated by the dotted lines. In contrast to the whole-cell configuration experiments, the current–voltage relationship for the main conductance of the human α4β2 nAChR is linear and shows no rectification (C). The amplitude values collected for six outside-out patches were fitted (straight line) with the current equation i = γ × (E − Er), where γ is the conductance, Er is the reversal potential, and E is the holding potential. The human α4β2 nAChR displays a main conductance of 46 pS with a reversal potential of −6.8 mV, confirming the high permeability of this ligand-gated channel for Na+ and K+ ions.X corresponds to single-channel amplitudes measured on a patch lasting long enough to allow examination of single-channel apertures at positive voltages. D, Current traces recorded at −100 and 60 mV. E, Loss of rectification in the outside-out configuration was also observed on large membrane patches containing numerous receptor proteins. Traces were recorded at −60 and 40 mV, respectively. In D–E, currents were elicited by 1 μm ACh (horizontal bar) applications.
The single-channel currents did not rectify when patches were held at positive potentials (Fig. 7C,D), as observed previously in other outside-out recordings of nAChRs (Ballivet et al., 1988; Mathie et al., 1990; Neuhaus and Cachelin, 1990; Mulle et al., 1992). The lack of rectification is further reinforced when recordings are made from outside-out patches containing a large number of channels (Fig.7E). Compared with the whole-cell recordings data (Fig. 6), currents recorded in these patches showed no rectification at positive potentials. The linear relationship observed between current and voltage indicates that conductance of these channels follows Ohm’s law.
As shown in Figure 8, ACh-evoked single channels in outside-out patches are characterized by a very fast rundown that can be fit by a single exponential process whose time constant is ∼47 sec. A similar rundown was observed in many patches. Its time course was independent of the ACh concentration or the application protocol and could not be prevented by adjunction of BAPTA, MgATP, or GTP in the patch pipette.

ACh-elicited single channels present fast run-down properties. In the outside-out configuration, the activity of the channels disappeared within minutes. The current recorded on a typical patch is presented in A, and the corresponding peak current amplitudes were plotted as a function of time inB. The current–time values were fitted with a single exponential function giving a time constant of 46.8 sec.
Effects of extracellular Ca2+ on the human α4β2 nAChR
It is known that extracellular Ca2+ potentiates, in a dose-dependent manner, the agonist-evoked whole-cell currents of native or reconstituted neuronal nAChRs (Mulle et al., 1992; Vernino et al., 1992; Eiselé et al., 1993) and is therefore considered a positive allosteric modulator (Léna and Changeux, 1993). To characterize the putative influence of extracellular Ca2+ on human α4β2 nAChRs, several experimental paradigms were designed. Determination of the current–voltage relationships in three external calcium concentrations (0.2, 2, 20 mm) revealed that the amplitude of the evoked currents was maximal at 2 mm (Fig.9A). Furthermore, these calcium effects were independent of the polarity of the ramp protocol (data not shown). Identical results were observed when the cell membrane was held at −100 mV (Fig. 9B). Although expected for low calcium concentrations, the reduction of the amplitude of the ACh-evoked currents observed for higher external calcium content suggests that several mechanisms might play a role in the modulation of the human α4β2 nAChR. To evaluate the kinetics and mechanism underlying the calcium effects, calcium jump experiments on steady-state ACh-evoked currents were performed (Fig. 9C). Currents recorded under these conditions revealed that fast and fully reversible inhibition of the nAChR-elicited current occurred.

High external Ca2+ decreases both whole-cell and single-channel currents of the human α4β2 nAChR. Voltage ramps (see Fig. 6 legend) were performed in external solutions containing (in mm): 130 NaCl, 10 HEPES, 20 glucose, and varying concentrations of CaCl2. When external calcium is increased from 0 to 2 mm, a significant potentiation of the ACh-evoked current for holding potentials less than −50 mV was observed; however, high calcium concentrations (10 or 20 mm) strongly inhibit the ACh-evoked current for holding potentials below 0 mV. A displays typical current–voltage relationships recorded on a single cell in three different calcium concentrations (1 μm ACh). For clarity, the current value is plotted once every 20 recorded points, corresponding to approximately one measurement every 7.5 mV. Continuous lines were computed using the same equation as for Figure 6. The calcium effects have been quantified by measuring for each cell the amplitude of the plateau currents (elicited by 1 μm ACh, 500 msec) recorded in three different calcium concentrations at −100 mV (B). The mean amplitude of the currents recorded in 0.2 mm calcium represents 47.8 ± 4.1% of the mean current measured in 2 mm calcium, whereas the mean current recorded in 20 mm calcium represents only 35.8 ± 4.3% (5 cells). High extracellular calcium is a negative modulator of the human α4β2 nAChR (C). When external Ca2+ jumped to 10 and 20 mm during the ACh pulse, a fast and fully reversible block of the ACh-evoked current was observed, indicating the concentration dependence of the calcium effect (n = 3). A high extracellular calcium concentration decreases the single-channel conductance of the human α4β2 nAChR (D–F). Holding potentials of −100, −80, −60, −40, −20, and 20 mV were changed at 5 sec intervals. One second after the voltage setting, the patch was challenged by 3 μm ACh for 200 msec. Such protocol was applied alternately in 20 and 2 mm CaCl2 (3 patches).D presents two traces of the records performed successively in 20 and 2 mm CaCl2. Even in the presence of the fast run-down mechanism, the total amount of current is much larger in 2 than in 20 mm Ca2+ (at all membrane potentials tested). The horizontal barrepresents the 3 μm ACh pulse. Reduction of the single-channel conductance induced by high extracellular calcium is illustrated in E. Patch was held throughout the experiment at −100 mV. Single-channel amplitudes (corresponding to the main conductance observed) were plotted as a function of the holding potential (F) and fit with the equationi = γ × (E −Er), where i is the current, γ is the conductance, E is the holding voltage, andEr is the current reversal potential. In 20 mm Ca2+, the main conductance decreases from 46 to 28 pS (see text).
To characterize the effect of high extracellular Ca2+concentrations on the single-channel currents at several potentials, protocols allowing investigation of the current over a broad voltage range in different calcium concentrations were designed. Data obtained from a single patch recorded successively in 20 and 2 mm of extracellular Ca2+ is presented in Figure 9D. The amount of current recorded in 2 mm Ca2+ was much larger (at all potentials) than equivalent currents measured in 20 mm Ca2+. The main unitary amplitude (as illustrated in Fig. 9E) was measured for three patches in both Ca2+ concentrations and plotted as a function of the holding potential (Fig. 9F). The conductance decreased from 46 to 28 pS when calcium was raised from 2 to 20 mm. Reversal potentials estimated by linear interpolation yielded intercepts ranging between −9 and −6 mV in 2 mmCa2+ and between −8 and −5 mV in 20 mmCa2+.
DISCUSSION
Functional expression of the human α4β2 nAChR
Reverse transcription-PCR analysis of total RNA isolated from the transfected (k177 clone), but not untransfected, cell line indicated the appropriate translation of mRNAs corresponding to α4 and β2 cDNAs (Gopalakrishnan et al., 1996). Moreover, ACh (1 mm) elicited no detectable current in wild-type HEK 293 cells, confirming the absence of functional nAChRs in the membranes of untransfected cells (Gopalakrishnan et al., 1995). Identical results were obtained in the absence of atropine. Thus, the ACh-evoked currents observed in K177 cells can be attributed only to the functional expression of the α4 and β2 subunits.
Pharmacological profile of the human α4β2 nAChR
Using a fast perfusion method (Puchacz et al., 1994;Gopalakrishnan et al., 1995; Bertrand et al., 1997) (Fig. 1), we have established that human α4β2 nAChRs display the following sensitivity: (−)-nicotine > ACh > (−)-cytisine > ABT-418. The small difference in apparent affinities between ACh and (−)-nicotine, however, remains to be confirmed by measurements on a larger sample of cells and different experimental protocols. When expressed in Xenopus laevis oocytes, human α4β2 nAChRs present an equivalent sensitivity and Hill coefficient for ACh (Sonia Bertrand, personal communication). Although ABT-418 is 10-fold less potent than (−)-nicotine in activating human α4β2 nAChRs, it is significantly more efficacious. Given the fast application method used in these experiments, and the relatively slow responses of the α4β2 nAChRs, the differences in the potency of ABT-418, (−)-nicotine, and ACh seem not to be attributable to partial desensitization of the receptor to compounds like (−)-nicotine or ACh. Thus it is possible that ABT-418 preferentially stabilizes the opened (and conducting) state(s) of the nAChRs and thereby evokes larger currents. It is noteworthy that ABT-418 behaves as a partial agonist on the human homomeric α7 nAChR expressed in HEK 293 cells where detectable currents were observed only for concentrations of ABT-418 above 300 μm (B. Buisson, J. P. Sullivan, and D. Bertrand, unpublished observations). These data illustrate the “pharmacological signature” displayed by different nAChRs subtypes within the same species.
In contrast to ABT-418, the anabaseine-derivative GTS-21, which is currently in clinical trial for the treatment of Alzheimer’s disease, behaves as a very weak agonist of the α4β2 nAChR. The amount of current elicited by GTS-21 was only 6% of the ACh-evoked currents (Fig. 3B,D). These data are in good agreement with those obtained previously with the rat α4β2 nAChRs reconstituted inXenopus oocyte (Hunter et al., 1994; De Fiebre et al., 1995). The observation that this anabaseine-derived compound acts as a stronger agonist of the rat α7 homo-oligomeric nAChR (De Fiebre et al., 1995) further reinforces the concept of a pharmacological signature that is specific to each neuronal nAChR.
Homomeric α7 nAChRs are blocked by MLA at nanomolar concentrations (Wonnacott et al., 1993; Gopalakrishnan et al., 1995; Palma et al., 1996), whereas the human α4β2 subtype is fully inhibited at concentrations above 10 μm (see Fig. 5).
Analysis of the potency of another nAChR antagonist such as DHβE reveals that an IC50 of 0.08 μm at the human α4β2 nAChRs compares rather well with the values obtained for the avian α4β2 nAChRs (Pereira et al., 1994) as well as the value determined for “type II” currents of cultured rat hippocampal neurons (Alkondon and Albuquerque, 1993). It was proposed that these currents result mainly from the activation of receptors comprising α4 and β2 subunits and that they were completely inhibited by infusion of 10 nm DHβE (Alkondon and Albuquerque, 1993). Similar preliminary results have been described with human nAChRs expressed inXenopus oocytes: DHβE inhibits much more selectively α4- and α2- than α3-containing receptors (Chavez-Noriega et al., 1995;Wong et al., 1995). Further investigations should confirm whether DHβE could be used as a relative selective inhibitor of α4- (and α2-) versus α3-containing nAChRs. Other experiments are needed, however, to assess the possible contribution of β subunits in the DHβE sensitivity (Duvoisin et al., 1989; Luetje et al., 1990; Hussy et al., 1994; Cachelin and Rust, 1995; Corringer et al., 1995; Harvey and Luetje, 1996).
Comparison of electrophysiology with86Rb+ efflux
In previous characterization of the human α4β2 nAChR (Gopalakrishnan et al., 1996), it was shown using rubidium (86Rb+) efflux that this receptor is about 10× more sensitive to (−)-nicotine than ACh. Comparison of these data with characterizations of the α4β2 nAChRs (Bertrand et al., 1990;McGehee and Role, 1995; for review, see Lindstrom, 1996) from other species suggested that human receptor might display a specific pharmacological profile. In contrast, however, data obtained with patch-clamp recordings (see above) are in good agreement with results from other studies (for review, see McGehee and Role, 1995). Thus, it seems that the initial discrepancies are attributable to the differences in techniques with flux measurements, on the one hand, and with electrophysiological recordings, on the other hand. The86Rb+ efflux technique requires long incubations (in the minute range) in the presence of the agonist, whereas millisecond exposures are typically used in electrophysiological recordings. Thus, the apparent discrepancies observed between these two experimental approaches might be attributed to inevitable desensitization of the receptor occurring during the ion flux protocol.
Rectification properties of the human α4β2 nAChR
Current–voltage relationships recorded in the whole-cell configuration either by voltage steps or by voltage ramps displayed a strong outward rectification (Fig. 6A,B). Additionally, rectification was not modified either by removal of intracellular Mg2+ or combined with the addition of the Mg2+ chelator CDTA in the pipette. In contrast, and independent of the presence of intracellular Mg2+, rectification was absent from the single-channel recordings obtained in the outside-out configuration (Fig. 8D,E; and see below). A similar observation was reported previously for the rat medial habenular neuron nAChRs (Mulle et al., 1992), a region known to express mRNAs encoding for α4 and β2 nAChR subunits (Mulle et al., 1991). The mechanisms underlying neuronal nAChR rectification have been analyzed more extensively in rat sympathetic ganglion neurons (Mathie et al., 1990) and in the PC12 cell line (Neuhaus and Cachelin, 1990;Ifune and Steinbach, 1991, 1992, 1993; Sands and Barish, 1992). Data obtained by Mathie et al. (1990) allowed them to conclude that in native heteromeric nAChRs, rectification cannot be attributed to the effect of intracellular Mg2+ alone. In contrast, however, rectification of both native and reconstituted α7 subunits can be abolished when Mg2+ is removed from the intracellular medium (Albuquerque et al., 1995; Forster and Bertrand, 1995;Bonfante-Cabarcas et al., 1996). These observations indicate that the mechanisms underlying rectification in α4β2 or α7 are distinct. Given the fast rundown and thus the absence of possible analysis of the opening probability or the burst duration, however, differences between nAChRs recorded in whole-cell or outside-out configurations cannot be ruled out in the present study. Thus, in agreement with Mathie et al. (1990), our data suggest that a factor other than Mg2+ must be invoked to explain the rectification of the human α4β2 nAChR observed in the whole-cell configuration. Recent findings indicate that intracellular polyamines determine the rectification of the potassium channel inward rectifier (Ficker et al., 1994; Lopatin et al., 1994;Falker et al., 1995) or of the kainate/AMPA subtypes of the glutamate receptors (Bowie and Mayer, 1995). Further experiments will be needed to investigate the possible effects of these compounds on native nAChRs.
High external Ca2+ inhibits human α4β2 nAChR whole-cell currents
Vernino et al. (1992) and Mulle et al. (1992) have shown that extracellular Ca2+ is a positive modulator of rat neuronal nAChRs (Léna and Changeux, 1993). In those studies, Ca2+ increased the whole-cell current amplitude while at the same time reducing the single-channel conductances, and these effects are more pronounced at lower agonist concentrations. In the present study, we observed that whole-cell currents are decreased both in low external Ca2+ (i.e., [Ca2+] <2 mm) and in high external Ca2+ (i.e., [Ca2+] >2 mm), with an optimal amplitude at ∼2 mm. This suggests that the positive Ca2+-induced modulation is observed for low Ca2+ concentrations (0.2–2 mm) but might be absent, or overcome by another mechanism, at higher concentrations. The inhibitory effect of high external Ca2+ differentiates the human α4β2 nAChR from the other characterized neuronal nAChRs (Mulle et al., 1992; Vernino et al., 1992). We observe a significant reduction of the single-channel conductance in a high calcium concentration (see Fig. 9), a result that is in full accordance with previous findings (Mulle et al., 1992; Vernino et al., 1992). The single-channel amplitude decrease could be interpreted as a screening mechanism for divalent cations (Imoto et al., 1988; Decker and Dani, 1990; Cooper et al., 1991). It is of value to note that a short pulse of 20 mm Ca2+ decreases the ACh-evoked current by ∼40% (Fig. 9C). Because this decrease is comparable to the reduction of the single-channel conductance (39%), it is probable that calcium inhibition results mainly from a screening charge effect. At present, however, involvement of other mechanisms cannot be ruled out.
In the absence of definitive assessment of the native neuronal nAChR stoichiometry and determination of the complete subunit family, one might postulate that an additional α (Conroy et al., 1992) or β subunit may be necessary for the reconstitution of the native human α4β2 containing nAChRs. Although this crucial question remains open at present, it is clear that the results described herein will facilitate studies probing the functional differences in α4β2 nAChRs that might be observed in human pathologies.
Footnotes
This research was supported by grants from the Swiss National Science Foundation (D.B.), the Office Federal de l’Education et des Sciences (D.B.), and the Human Frontier Science Program (D.B.). B.B. is the recipient of a research fellowship from the J. Thorn Foundation. We thank Sonia Bertrand for comments and suggestions during the preparation of this manuscript.
Correspondence should be addressed to Daniel Bertrand, Department of Physiology, University of Geneva, 1 Rue M. Servet, CH-1211 Geneva 4, Switzerland.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}