


Abstract
Nitric oxide (NO) plays an important role in inflammation and multiple stages of carcinogenesis. We investigated the effect of various tea polyphenols and caffeine on the induction of NO synthase (NOS) in thioglycollate-elicited and lipopolysaccharide (LPS)-activated peritoneal macrophages. Gallic acid (GA), (−)-epigallocatechin (EGC), and (−)-epigallocatechin-3-gallate (EGCG), the major tea catechin, were found to inhibit inducible NOS (iNOS) protein in activated macrophages. EGCG, a potent antitumor agent with anti-inflammatory and antioxidant properties, inhibited NO generation, as measured by the amount of nitrite released into the culture medium. Inhibition of NO production was observed when cells were cotreated with EGCG and LPS. iNOS activity in soluble extracts of lipopolysaccharide-activated macrophages treated with EGCG (5 and 10 μm) for 6–24 hr was significantly lower than that in macrophages without EGCG treatment. Western blot, reverse transcription-polymerase chain reaction, and Northern blot analyses demonstrated that significantly reduced 130-kDa protein and 4.5-kb mRNA levels of iNOS were expressed in lipopolysaccharide-activated macrophages with EGCG compared with those without EGCG. Electrophoretic mobility shift assay indicated that EGCG blocked the activation of nuclear factor-κB, a transcription factor necessary for iNOS induction. EGCG also blocked disappearance of inhibitor κB from cytosolic fraction. These results suggest that EGCG decreases the activity and protein levels of iNOS by reducing the expression of iNOS mRNA and the reduction could occur through prevention of the binding of nuclear factor-κB to the iNOS promoter, thereby inhibiting the induction of iNOS transcription.
NO is a short-lived bioactive molecule that participates in the physiology and pathophysiology of many systems (1). NO is synthesized in vivo from l-arginine by NOS with NADPH and oxygen as cosubstrates (2). Molecular cloning and sequencing analyses revealed the existence of at least three main types of NOS isoforms. NOS in the endothelium of blood vessels and brain neurons is constitutive and Ca2+/calmodulin dependent. This constitutive enzyme synthesizes small amounts of NO triggered by various agonists (e.g., bradykinin, acetylcholine) that increase intracellular Ca2+. NOS in macrophages and hepatocytes is inducible, not detectable, in unstimulated cells and requires protein synthesis for expression. iNOS produces large amounts of NO several hours after exposure to endotoxin and/or cytokines in macrophages, Kupffer cells, hepatocytes, and fibroblasts. NO has a wide biological role in modulating physiological and pathophysiological processes (1, 3), such as macrophage cytotoxicities, neurotransmissions, neurotoxicities, and regulation of blood pressure. Low concentrations of NO are sufficient, in most cases, to effect these functions. However, during infection and inflammation, in vivo formation of NO is increased, suggesting that NO concentrations are well above those found in normal physiological function in infected tissues. High concentrations of NO have been shown to cause deamination of deoxynucleotides and bases within intact DNA in vitro and are mutagenic in vivo (4). Exposure of human cells to NO under aerobic conditions results in DNA strand breakage and nitrosative deamination of DNA bases (5). Therefore, NO can cause DNA damage as well as mutation in human cells.
A portion of the 5′-flanking region of the murine iNOS gene has been cloned (6). The promoter of the murine gene contains a TATA box and consensus sequences for the binding of transcription factors associated with stimuli that induce iNOS expression (7). These potentially relevant transcription factors, NF-κB, and interferon regulatory factor, have been shown to be functionally important for iNOS induction.
Tea is one of the most popular beverages in the world because of its attractive flavor and aroma. Polyphenols are the most significant group of tea components, especially the catechin group of the flavonols. The major tea catechins are EGCG, EGC, ECG, EC, (+)-gallocatechin, and (+)-catechin. Many biological functions of tea polyphenols have been studied (8), including anti-inflammatory, antioxidative (9-11), antimutagenic (12), and anticarcinogenic (13) effects. Biochemical mechanisms for inhibition of tumorigenesis by tea might occur via five mechanisms: (a) antioxidative function, (b) inhibition of nitrosation reactions, (c) modulation of xenobiotic metabolizing enzymes, (d) trapping of activated forms of carcinogens, and (e) inhibition of activities related to tumor promotion and cell proliferation. The anti-inflammatory and cancer-preventative characteristics of EGCG have been well documented. How these inhibitory responses are modulated by EGCG is not well characterized.
In the current study, we examined the effects of EGCG on NO production from murine peritoneal macrophages and sought possible reaction mechanisms. The data suggest that EGCG blocks early events of NOS induction, and the blocking could occur via inhibition of binding of transcription factor NF-κB to the iNOS promoter, thereby inhibiting the induction of iNOS transcription.
Materials and Methods
Reagents.
LPS (Escherichia coli 0127:B8), GA, sufanilamide, naphthylethylenediamine dihydrochloride, DTT, NADPH, FAD, and FMN were purchased from Sigma Chemical (St. Louis, MO). Caffeine was purchased from Merck (Darmstadt, Germany). Catechin and EC were purchased from Aldrich Chemical (Milwaukee, WI). Isotopes were obtained from Amersham (Arlington Heights, IL). Polynucleotide kinase and oligo(dT)18 were obtained from Pharmacia (Piscataway, NJ). EGCG, EGC, and EC were purified as previously described (9); their purities were 97%, 95%, and 93%, respectively.
Cell culture.
Thioglycollate-elicited peritoneal macrophages were obtained from specific pathogen-free male BALB/c mice at 5–8 weeks of age by injection of 3.0 ml of sterile 3% thioglycollate solution (Difco, Detroit, MI) for 4 days before lavage with 10 ml of phosphate-buffered saline. The PECs were centrifuged at 1500 rpm and washed once with RPMI-1640 (without phenol red) supplemented with 10% endotoxin-free heat-inactivated fetal calf serum (GIBCO, Grand Island, NY), 100 units/ml penicillin, and 100 μg/ml streptomycin. The cells were resuspended in RPMI-1640 at a density of 2 × 106cells/ml. Viability was >95% as determined by trypan blue dye exclusion. The PECs were plated and incubated for 2–3 hr at 37° in humidified 5% CO2/95% air to allow macrophage adherence. The plates were then washed once with warm RPMI-1640 to remove nonadherent cells. This procedure resulted in an adherent cell population that was 85–95% macrophages, as determined by morphology and esterase staining.
Nitrite assay.
The nitrite concentration in the culture medium was measured as an indicator of NO production according to the Griess reaction (14). One hundred microliters of each supernatant was mixed with the same volume of Griess reagent (1% sulfanilamide in 5% phosphoric acid and 0.1% naphthylethylenediamine dihydrochloride in water); absorbance of the mixture at 550 nm was determined with an enzyme-linked immunosorbent assay plate reader (Dynatech MR-7000; Dynatech Labs, Chantilly, VA).
Determination of NOS activity.
The iNOS specific activity was determined by [3H]arginine-to-[3H]citrulline conversion assay (15). Macrophage lysate was incubated with 50 mmHEPES, pH 7.4, 15 nm [3H]arginine, 1 mm NADPH, 1 mm EDTA, 1.25 mmCaCl2, 1 μg of calmodulin, 5 μm FAD, 5 μm FMN, 0.1 mm tetrahydrobiopterin, 1 mm DTT, inhibitors of arginase, and other urea cycle enzymes (50 mm valine, 1 mm ornithine, 1 mm citrulline) in the presence and absence of 1 mm N G-monomethyl-l-arginine for 15 min at 37°. The NOS activity was determined from the difference between the [3H]citrulline produced with and withoutN G-monomethyl-l-arginine. The protein concentration was determined with bicinchoninic acid protein assay (Pierce, Rockford, IL).
Western blots.
Total cellular extract was prepared using radioimmunoprecipitation assay buffer (50 mm Tris·HCl, pH 7.4, 150 mm NaCl, 1% Triton X-100, 1% deoxycholate, 0.1% sodium dodecyl sulfate, 1% aprotinin). Total protein (for iNOS) or cytosolic fractions (for IκB) containing 50 μg of protein were separated on sodium dodecyl sulfate-polyacrylamide minigels (8% for iNOS and 10% for IκB) and transferred to Immobilon polyvinylidene difluoride membranes (Millipore, Bedford, MA). The membrane was incubated overnight at 37° with 10% bovine serum albumin in phosphate-buffered saline to block nonspecific immunoglobulins and then incubated with anti-macNOS monoclonal antibody (Transduction Laboratories, Lexington, KY), anti-IκBα polyclonal antibody (Santa Cruz Biochemicals, Santa Cruz, CA), or anti-α-tubulin monoclonal antibody (Oncogene Science, Cambridge, UK). iNOS, IκB, and α-tubulin protein were detected by chemiluminescence (ECL, Amersham).
RT-PCR.
Total RNA was isolated by acid guanidinium thiocyanate-phenol-chloroform extraction (16). Total RNA (1 μg) was converted to cDNA with 1 μm oligo(dT)18, 0.5 mm concentration of each dNTP, 50 mmTris·HCl, pH 8.3, 75 mm KCl, 3 mmMgCl2, 1 unit/μl RNase inhibitor, and 10 units/μl Moloney murine leukemia virus reverse transcriptase at 42° for 1 hr. The amplification of iNOS cDNA was performed by incubating 20-ng equivalents of DNA in 100 mm Tris·HCl buffer, pH 8.3, containing 500 mm KCl, 15 mm MgCl2, 0.1% gelatin, 200 μm concentration of each dNTP, and 50 units/ml Super Taq DNA polymerase with the following oligonucleotide primers: 5′-CCCTTCCGAAGTTTCTGGCAGCAGC-3′ and 5′-GGCTGTCAGAGAGCCTCGTGGCTTTGG-3′. The cDNA sequence of glyceraldehyde-3-phosphate dehydrogenase was also amplified as control in a similar way using the following primers: 5′-TGAAGGTCGGTGTGAACGGATTTGGC-3′ and 5′-CATGTAGGCCATGAGGTCCACCAC-3′. A thermal cycle of 45 sec at 95°, 45 sec at 65°, and 2 min at 72° was used for 30 cycles. PCR products were analyzed on 1.8% agarose gels (15).
Preparation of probe.
PCR primers for mouse iNOS were synthesized according to the following oligonucleotide sequences (17): forward primer, 5′-CCCTTCCGAAGTTTCTGGCAGCAGC-3′, 2944–2968; and reverse primer, 5′-GGCTGTCAGAGAGCCTCGTGGCTTTGG-3′, 3416–3440. Total cellular RNA (5 μg) from LPS-stimulated macrophages of BALB/c mice was used as a template, and the single-stranded cDNA was synthesized with oligo(dT)18 by reverse transcriptase. cDNA was amplified by using a DNA thermal cycler Perkin-Elmer Cetus (Norwalk, CT) in 35 cycles of denaturing at 95° for 45 sec, annealing at 65° for 45 sec, and extension at 72° for 2 min. The PCR product of expected size (497 bp) was ligated into the pGEM plasmid vector (Promega, Madison, WI) by the T/A cloning method (18). Nucleotide sequence of the subcloned iNOS cDNA was determined by the dideoxy chain termination method (19) (USB DNA sequencing kit; Amersham Life Science, Clearbrook, IL) and coincided with that of iNOS as previously reported (17).BamHI/EcoRI restriction fragments of the subcloned cDNA for iNOS was used as probe for Northern blot analysis.
Northern blot analysis.
Total RNA (20 μg) were separated by electrophoresis on a 1.2% agarose gel containing 6.7% formaldehyde and transferred to a Hybond-N nylon membrane (Amersham Life Science) in 20× standard saline citrate (3 m sodium chloride and 0.3m sodium citrate, pH 7.0). After baking at 80° for 2 hr and prehybridization for 4 hr, the filters were hybridized with32P-labeled cDNA probes at a concentration of 3 × 106 cpm/ml for 16–18 hr at 42°. The probe was labeled with [α-32P]dCTP by using a Random Primer labeling kit (Stratagene, La Jolla, CA). Then, the filters were washed, dried, and autoradiographed with Kodak X-Omat XAR-film using intensifying screens at −80°. After hybridization with iNOS-specific probe, the blot was stripped and reprobed with a probe for GAPDH cDNA (20) as a control.
Preparation of extracts and electrophoretic mobility shift assay.
Nuclear and cytoplasmic extracts were prepared according to a modified method of Chen et al. (21). At the end of culture, the cells were suspended in hypotonic buffer A (10 mm HEPES, pH 7.6, 10 mm KCl, 0.1 mmEDTA, 1 mm DTT, 0.5 mm phenylmethylsulfonyl fluoride) for 10 min on ice and vortexed for 10 sec. Nuclei were pelleted by centrifugation at 12,000 × g for 20 sec. The supernatants containing cytosolic proteins were collected. A pellet containing nuclei was resuspended in buffer C (20 mm HEPES, pH 7.6, 25% glycerol, 0.4 m NaCl, 1 mm EDTA, 1 mm DTT, 0.5 mm phenylmethylsulfonyl fluoride) for 30 min on ice. The supernatants containing nuclear proteins were collected by centrifugation at 12,000 × g for 2 min and stored at −70°. For electrophoretic mobility shift assay, each 10 μg of nuclear extract was mixed with the labeled double-stranded NF-κB oligonucleotide, 5′-AGTTGAGGGGACTTTCCCAGGC-3′, and incubated at room temperature for 20 min (underlining indicates κB consensus sequence or binding site for NF-κB/c-Rel homodimeric and hetero dimeric complexes). The incubation mixture included 1 μg of poly(dI-dC) in a binding buffer (25 mm HEPES, pH 7.9, 0.5 mm EDTA, 0.5 mm DTT, 1% Nonidet P-40, 5% glycerol, 50 mm NaCl). The DNA/protein complex was electrophoresed on 4.5% nondenaturing polyacrylamide gels in 0.5× Tris/borate/EDTA buffer (0.0445 m Tris, 0.0445m borate, 0.001 m EDTA). A double-stranded mutated oligonucleotide, 5′-AGTTGAGGCGACTTTCCCAGGC-3′, was used to examine the specificity of binding of NF-κB to DNA (the underlined sequence is identical to κB consensus sequence except for a G-to-C substitution in the NF-κB/Rel DNA binding motif). The specificity of binding was also examined by competition with the unlabeled oligonucleotide.
Results
EGCG inhibits NO generation.
The inhibition of NO generation in the LPS-activated macrophages with EGCG was time dependent (Fig.1). Maximum inhibition was observed when thioglycollate-elicited peritoneal macrophages were cotreated with EGCG (10 μm) and LPS (5 μg/ml). However, when EGCG was added 3, 6, or 18 hr after activation or 1 or 2 hr before activation, with or without washing of EGCG, the inhibition of NO generation was less effective. Therefore, in this study we used cotreatment of macrophages with LPS and EGCG in most experiments.

Effect of EGCG (10 μm) pretreatment, cotreatment, or post-treatment of macrophages activated with LPS (5 μg/ml). Thioglycollate-elicited peritoneal macrophages were cultured for 24 hr in the presence of EGCG (10 μm) before (−2 hr, −2 hr + wash, −1 hr, −1 hr + wash), during (0 hr), or after (+3 hr, +6 hr,+18 hr) activation of macrophages with LPS.
EGCG inhibits iNOS activity.
Arginine-to-citrulline conversion assays were performed to measure the activity of iNOS in macrophages. iNOS activity in soluble extracts of activated macrophages in the absence of EGCG was increased in a time-dependent manner (within 6–24 hr). In the presence of EGCG (5 and 10 μm), the specific activity of iNOS was significantly inhibited in a dose-dependent manner (Fig. 2). The presence of EGCG (10 μm) inhibited induction of iNOS activity by 55.1% at 24 hr.

NOS activity in activated macrophages with and without EGCG. Results are the mean values of triplicate assays. Similar results were obtained in three other experiments.
EGCG inhibits iNOS protein.
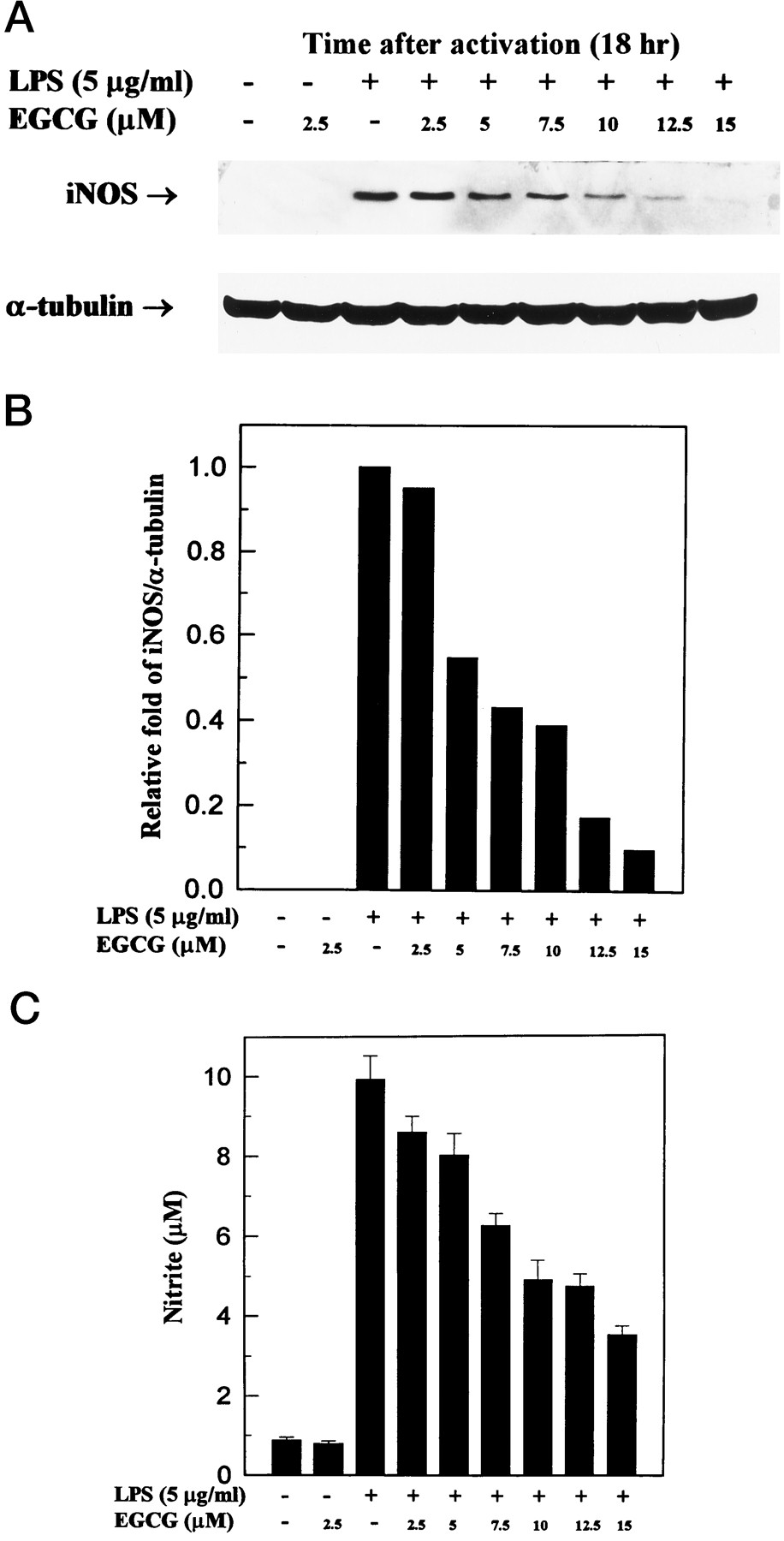
The dose response of EGCG on the inhibition of 130-kDa iNOS protein and NO generation is shown in Fig.3. At 2.5, 5, 7.5, 10, 12.5, and 15 μm, EGCG inhibited the levels of iNOS protein by 5%, 45%, 57%, 61%, 83%, and 90%, respectively, and inhibited NO generation by 13%, 19%, 37%, 51%, 52%, and 65%, respectively. We also determined the time course of changes in synthesized iNOS protein and released NO (Fig. 4); st 6, 18, and 24 hr, the amounts of iNOS protein and NO were significantly lower in cells treated with 10 μm EGCG than in those not treated with EGCG.

Dose response of inhibition of LPS-dependent iNOS activation by EGCG. Western blotting analysis of iNOS (A) and nitrite release (C) in culture medium of activated macrophages with or without EGCG. Thioglycollate-elicited peritoneal macrophages were incubated with LPS (5 μg/ml) and various concentrations of EGCG for 18 hr. B, Band intensities were quantified by densitometry (IS-1000 Digital Imaging System). This experiment was repeated three times with similar results.
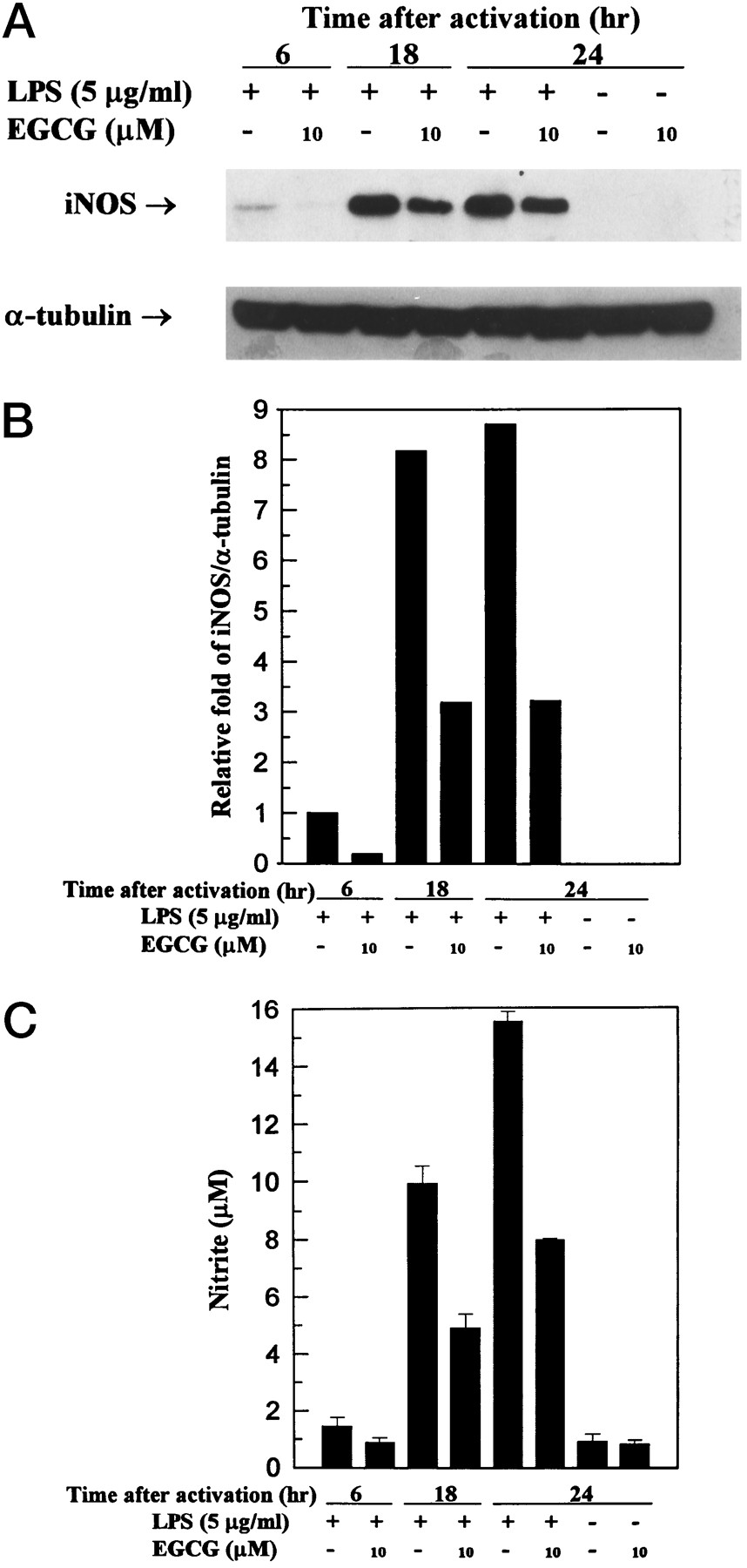
Effect of EGCG on LPS-induced iNOS activation at 6, 18, and 24 hr. Thioglycollate-elicited peritoneal macrophages were cultured in the presence of LPS (5 μg/ml) with or without EGCG (10 μm) for 6, 18, and 24 hr. At the end of the incubation time, the culture medium was collected for nitrite assay (C) and extraction of total protein for iNOS protein and α-tubulin analysis (A). B, Band intensities were quantified by densitometry (IS-1000 Digital Imaging System). This experiment was repeated three times with similar results.
EGCG inhibits iNOS expression.
Macrophages were activated with LPS with and without EGCG (10 μm) for 3 and 6 hr, harvested, and assayed for iNOS mRNA expression by RT-PCR (Fig.5). We found that significantly lower levels of iNOS mRNA were expressed in macrophages activated by LPS in the presence of EGCG than in its absence. These experiments were repeated three times with similar results. The data in Fig.6 demonstrate up-regulation of iNOS mRNA levels on stimulation with LPS for 3 and 6 hr. iNOS mRNA was present as a single band of ∼4.5 kb. In unstimulated cells, there was no detectable iNOS mRNA. The addition of EGCG alone had no effect on iNOS mRNA levels. Coincubation of macrophages with LPS plus EGCG caused a complete suppression of iNOS induction after 3-hr incubation, and after 6-hr incubation, only a weak induction of iNOS mRNA was detectable in the presence of EGCG.

RT-PCR analysis of mRNA expression of iNOS. A, Thioglycollate-elicited peritoneal macrophages were incubated in the presence of LPS (5 μg/ml) with or without EGCG (10 μm) for 3 and 6 hr. B, Band intensities were quantified by densitometry. This experiment was repeated three times with similar results.G3PDH, glyceraldehyde-3-phosphate dehydrogenase.
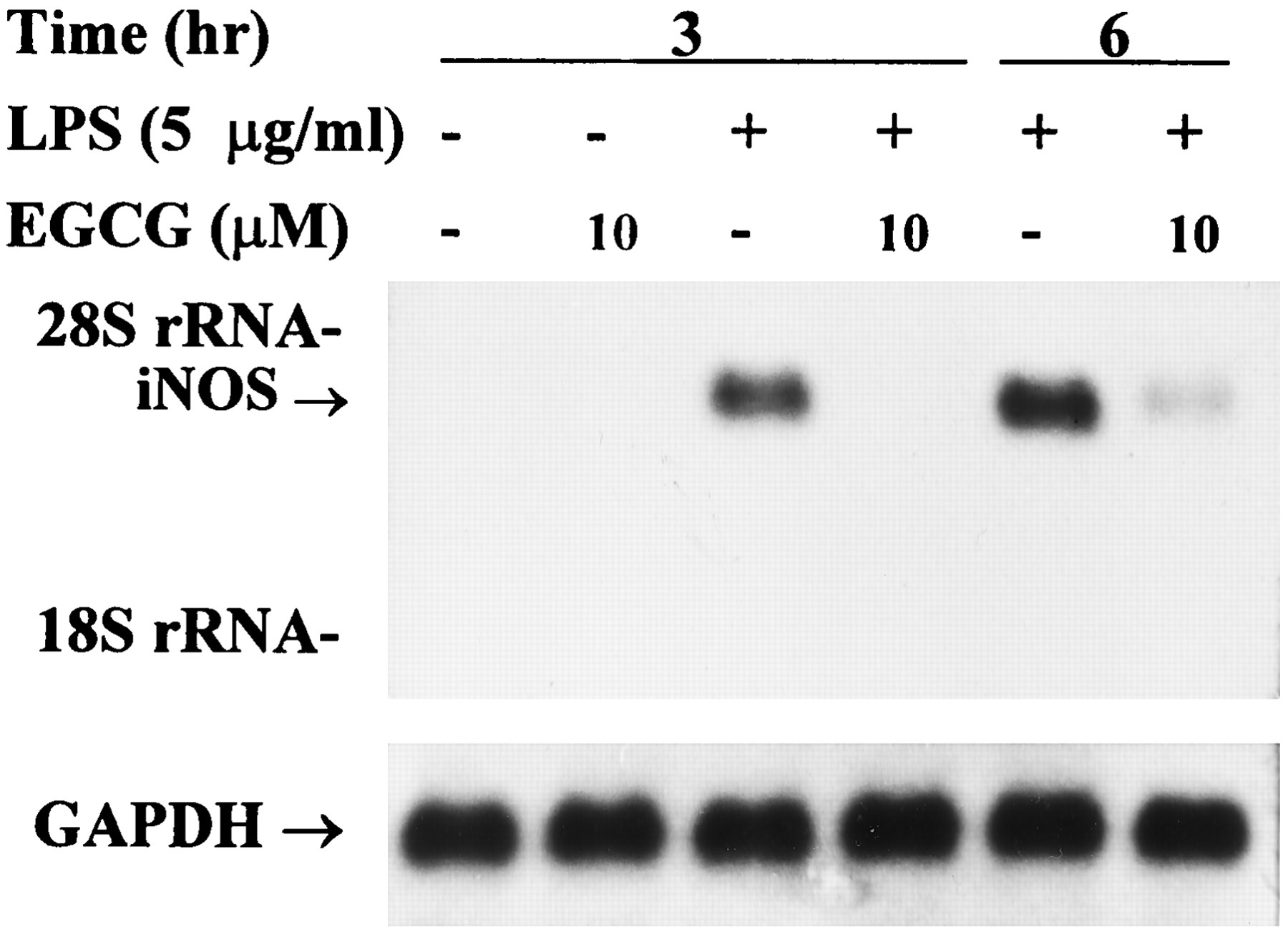
Northern blot analysis of mRNA expression of iNOS. Thioglycollate-elicited peritoneal macrophages were incubated in the presence of LPS (5 μg/ml) with or without EGCG (10 μm) for 3 and 6 hr. Total RNA (20 μg) was successively hybridized to32P-labeled iNOS and GAPDH probes as described in Materials and Methods. Arrows, 4.5-kb iNOS mRNA.Bottom, signals for GAPDH mRNA for each lane are shown as control. rRNA, ribosomal RNA.
Effects of various tea polyphenols and caffeine on iNOS protein.
Various tea polyphenols and caffeine were examined to determine whether they affect iNOS protein and NO production in macrophages activated with LPS for 18 hr (Fig.7). GA, EGC, EGCG, and EC were found to significantly reduce the amount of iNOS protein; their inhibition percentages were 92%, 74%, 61%, and 50% at a dose of 10 μm, respectively, whereas ECG, (+)-catechin, and caffeine showed less inhibitory effect at the same dose.

Inhibition of NOS induction in activated macrophages by various tea polyphenols and caffeine. Thioglycollate-elicited peritoneal macrophages were treated with LPS (5 μg/ml) and various tea polyphenols or caffeine (10 μm) for 18 hr. At the end of the incubation time, the culture medium was collected for nitrite assay (C) and extraction of total protein for iNOS protein and α-tubulin analysis (A). B, Band intensities were quantified by densitometry. C, (+)-catechin. This experiment was repeated three times with similar results.
Effect of EGCG on LPS-induced nuclear proteins with NF-κB binding activity.
Electrophoretic mobility shift assay using a consensus oligonucleotide for NF-κB binding showed a low level of NF-κB binding activity in untreated peritoneal macrophages. Incubation of peritoneal macrophages with LPS (5 μg/ml) for 3 hr markedly increased this activity. The induction of specific NF-κB binding with NF-κB site by LPS was markedly inhibited by coincubation with EGCG (10 and 15 μm) (Fig. 8a). The addition of excess unlabeled consensus oligonucleotide completely prevented the band shifts, demonstrating the specificity of the protein/DNA interaction. The addition of excess unlabeled mutated oligonucleotide did not prevent the band shifts, also demonstrating the specificity of the protein/DNA interaction (Fig. 8b).
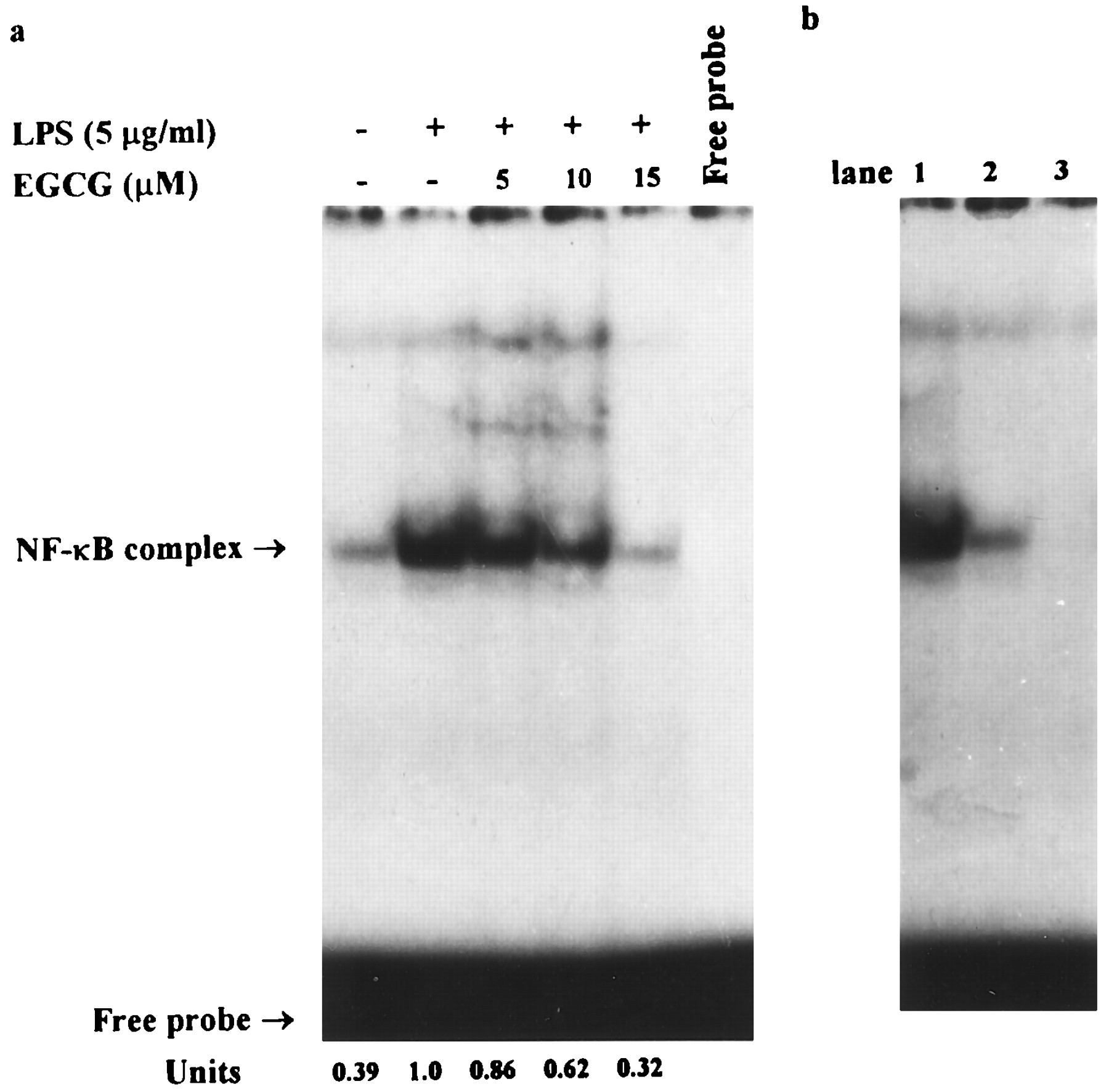
Electrophoretic mobility shift assay using a 5′-end-labeled consensus oligonucleotide for NF-κB binding and nuclear extracts from murine peritoneal macrophages. A, Mouse peritoneal macrophages were cotreated with LPS (5 μg/ml) without or with different concentrations (5–15 μm) of EGCG and then incubated for 3 hr. B, Specificity of NF-κB complex formation was verified in LPS-only sample by displacement with a 50-fold excess of the unlabeled mutated oligonucleotide (lane 1) and a 25- and 50-fold excess of the unlabeled consensus oligonucleotide (lanes 2 and 3). Band intensities were quantified by densitometry. The arbitrary units represent the relative amounts of the radioactivity present in respective bands. This experiment was repeated three times with similar results.
Effects of EGCG on degradation of IκB.
To determine whether the inhibitory action of EGCG was due to its effect on IκB degradation, the cytoplasmic levels of IκB protein were examined by Western blot analysis. When thioglycollate-elicited peritoneal macrophages were activated with LPS, IκB disappeared from cytosolic fractions. EGCG treatment at the same time of activation abolished the LPS-induced IκB degradation (Fig. 9). These results suggest that inhibition of NO production by EGCG occurred via the prevention of NF-κB activation.

LPS-mediated IκB degradation is blocked by the EGCG. Mouse peritoneal macrophages were cotreated with LPS (5 μg/ml) without or with different concentrations (5–15 μm) of EGCG and then incubated for 3 hr. Cytosolic fractions were prepared and analyzed for the content of IκB protein by Western blotting.
Discussion
EGCG is a biochemically active compound with known anti-inflammatory, anticarcinogenic, and free radical-scavenging properties. However, how EGCG carries out these functions is not very clear. We demonstrated that EGCG, EGC, and GA inhibit induction of iNOS in murine peritoneal macrophages activated with LPS. The results indicated that the galloyl and hydroxyl groups at the 3′ position on EGCG were responsible for its strongly anti-inflammatory property. These tea polyphenols have phenol rings that act as electron traps to scavenge peroxy radicals, superoxide anions, and hydroxyl radicals and prevent oxidation of iron (8-11). Therefore, we suggest that in addition to the reduction of iNOS expression, these compounds may block peroxynitrite and nitrite production through inhibition of oxidative reactions.
Mammals are in permanent contact with Gram-negative bacteria and LPS (22). Low doses of LPS are thought to be beneficial for the host (e.g., in causing immunostimulation and enhanced resistance to infections and malignancies). On the other hand, the presence of a large amount of LPS leads to dramatic pathophysiological reactions such as fever, leukopenia, tachycardia, tachypnea, hypotension, disseminated intravascular coagulation, and multiorgan failure. LPS stimulates host cells (mainly monocytes/macrophages but also endothelial cells, smooth muscle cells, and neutrophils) to produce and release endogenous mediators, such as NO. There are several mechanisms by which intracellular elevated NO can exert genotoxic affects after reacting with oxygen; these include formation of carcinogenicN-nitroso compounds, direct deamination of DNA bases (4,23), DNA strand breakage, and oxidation of DNA after formation of peroxynitrite and/or hydroxyl radicals. The iNOS isoform can produce high, persistent concentrations of NO on induction with endotoxin alone or in combination with cytokines in many cell types and is expressed in the resting state in other cells, potentially resulting in cytotoxicity, tissue damage, or DNA damage. NO reacts with oxygen to form NO2, which dimerizes to N2O4. N2O4 dismutates spontaneously in water to form nitrates and nitrites (2). Nitrate can be microbiologically reduced to nitrite (24), which can then interact with dietary substrates such as amines or amides to produce N-nitroso compounds. The formation of carcinogenic N-nitrosamines resulting from elevated NO formation has been demonstrated in cell cultures andin vivo (25); therefore, EGCG inhibition of iNOS activity or expression may block the formation of N-nitroso compounds and peroxynitrite or hydroxyl radicals and could thus inhibit carcinogenesis. EGCG may be internalized and reacts with ROIs.
EGCG decreased the activity and protein levels of iNOS by reducing the expression of iNOS mRNA. At the gene level, the expression of murine macrophage NOS is largely regulated by transcriptional activation. The promoter of the iNOS gene contains two major discrete regions synergistically functioning for binding of transcription factors (26-29): one for NF-κB, which is mainly activated by LPS, and the other one for interferon-related transcription factors, such as interferon regulatory factor-1. NF-κB is a mammalian transcription factor that controls a number of genes that are important for immunity and inflammation. NF-κB is composed mainly of two proteins: p50 and p65. In its unstimulated form, NF-κB is present in the cytosol bound to the inhibitory protein IκB. After induction of cells by a variety of agents, IκB becomes phosphorylated and triggers a proteolytic degradation of IκB; then, NF-κB is released from IκB and translocated to the nucleus. In 1996, Imbert et al. (30) reported an alternative mechanism of NF-κB activation. Stimulation of Jurkat T cells with the protein tyrosine phosphatase inhibitor and T cell activator pervanadate led to NF-κB activation through tyrosine phosphorylation but not degradation of IκB. Agents that have been described as NF-κB activators include mitogens, cytokines, LPS, 12-O-tetradecanoylphorbol-13-acetate, and cAMP. ROIs have also been proposed to be involved in the activation of NF-κB (28); this is based on the observations that treatment of some cells with H2O2 can activate NF-κB. EGCG has been demonstrated to be an antioxidant (9, 10); we found that EGCG can block activation of NF-κB. This is consistent with the observation that certain antioxidants, such as N-acetyl cysteine or pyrrolidine dithiocarbamate, can block activation of NF-κB by blocking the signal-induced phosphorylation of IκB.
We demonstrated that EGCG inhibits the binding of transcription factor NF-κB to the NF-κB binding site of the iNOS promoter in peritoneal macrophages, thereby inhibiting the induction of iNOS transcription. A similar finding was made by Kleinert et al. (31); they observed that dexamethasone decreased the activity of the iNOS promoter and reduced the formation of cytokine-induced NF-κB complexes that bind to the NF-κB site in the human iNOS promoter. They suggested that the dexamethasone-activated glucocorticoid receptor is likely to interact with the cytokine-activated NF-κB complex, thereby repressing the binding of this complex to the NF-κB response element in the 5′-flanking sequence of the iNOS gene.
Kleinert et al. (32) reported that in 3T3 cells, at least three different signal transduction pathways can stimulate iNOS mRNA expression: the receptor tyrosine kinase pathway (by interferon-γ, tumor necrosis factor-α, and LPS), the protein kinase A pathway (by forskolin, 8-bromo-cAMP, and 3-isobutyl-1-methylxanthine), and the protein kinase C pathway (by 12-O-tetradecanoylphorbol-13-acetate). All these pathways seem to converge in the activation of the essential transcription factor NF-κB. However, there is a marked intercell variability for the protein kinase A and C pathways. For example, in murine RAW 264.7 cells, neither the protein kinase A nor the protein kinase C pathway is able to stimulate this transcription factor; they even inhibit NF-κB-dependent reporter gene expression in response to LPS (33). In human Jurkat T cells, the protein kinase C pathway, but not the protein kinase A pathway, activates NF-κB (34). Conversely, in murine J774 macrophages, activators of protein kinase A are effective stimulators of NF-κB, whereas protein kinase C activators failed to stimulate this transcription factor (35). Thus, the NF-κB activation mechanisms seem to be cell specific. Kleinert et al. (32) also found that the phosphodiesterase inhibitor 3-isobutyl-1-methylxanthine produced a marked induction of iNOS mRNA in 3T3 cells. We demonstrated that although cotreatment of PEC with the phosphodiesterase inhibitor caffeine and LPS had no synergistic effect on iNOS production (Fig. 6), there was little efficacy in inhibition of iNOS. Shi et al.(36) demonstrated that caffeine effectively scavenges hydroxy radical (HO·) (36). Therefore, this effect of caffeine can be explained by the scavenging of HO· and then inhibition of iNOS protein and nitrite production. The increases in cAMP and cGMP by caffeine are unable to activate iNOS in PEC, but this requires further studies.
Phosphorylation of proteins seems to play a significant role in LPS signaling pathways, including protein tyrosine kinases, mitogen-activated protein kinase, protein kinase C, G protein, protein kinase A, and ceramide-activated protein kinase (37). LPS activates protein kinase C, which in turn induces NADPH oxidase activity (38). NADPH oxidase is located in the plasma membrane and uses intracellular NADPH to produce extracellular O2·−. O2·− is unstable and rapidly dismutes (within milliseconds) to form H2O2. H2O2 can rapidly cross cell membranes, whereas other ROIs, such as O2·− and HO·, cannot. Once inside the cell, H2O2 can react with iron or copper according to the Fenton reaction to form HO·. In our recent report, we found that HO· radical is rapidly generated after H2O2 enters into the cell; then, HO· can activate protein tyrosine kinase (39). The role of protein tyrosine kinase has also been implicated in NF-κB activation by LPS, ultraviolet, and hypoxia (38). However, we postulated that EGCG was likely through inhibition of LPS-induced phosphorylation and degradation of IκB. The step in the signal transduction pathway of NF-κB activation inhibited by EGCG is at or before the phosphorylation step of NF-κB. EGCG also has been shown to inhibit both protein kinase C and protein kinase A (40). Based on our recent results, we think that EGCG can significantly inhibit epidermal growth factor interaction with EGF receptor,1 and preincubation of EGCG with LPS could slightly repress LPS induction of iNOS protein (data not shown). Therefore, we also postulated that EGCG could block the interaction of LPS with its receptor, creating a “sealing effect.” EGCG may interact with LPS or its receptor and then block the LPS signal transduction pathway of NF-κB activation.
On the basis of the current results and those of other reports, we hypothesize that the possible site of action of EGCG on LPS-induced iNOS activation is as shown in Fig. 10. We conclude that EGCG prevents binding of the essential transcription factor NF-κB to the iNOS promoter, which in turn leads to decreased transcription of the iNOS gene. The signal transduction pathway of NF-κB activation inhibited by EGCG may be mediated through (1) inhibition of LPS interaction with its receptor, (2) scavenging ROIs, and (3) inhibition of protein kinases.

The possible site of action of EGCG on LPS-induced iNOS activation.
Footnotes
- Received December 26, 1996.
- Accepted April 16, 1997.
-
Send reprint requests to: Dr. Jen-Kun Lin, Institute of Biochemistry, College of Medicine, National Taiwan University, No. 1, Section 1, Jen-Ai Road, Taipei, Taiwan 100, R.O.C.
-
↵1 Y.-L. Lin and J.-K. Lin, unpublished observations.
-
This study was supported by National Science Council Grant NSC 86–2314-B-002–042 and National Health Research Institute Grant DOH 86-HR-403.
Abbreviations
- NO
- nitric oxide
- iNOS
- inducible nitric oxide synthase
- LPS
- lipopolysaccharide
- GA
- gallic acid
- EGC
- (−)-epigallocatechin
- EGCG
- (−)-epigallocatechin-3-gallate
- NF-κB
- nuclear factor-κB
- EC
- (−)-epicatechin
- ECG
- (−)-epicatechin-3-gallate
- ROI
- reactive oxygen intermediate
- PEC
- peritoneal exudate cell
- HEPES
- 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
- DTT
- dithiothreitol
- GAPDH
- glyceraldehyde-3-phosphate dehydrogenase
- PCR
- polymerase chain reaction
- RT
- reverse transcription
- IκB
- inhibitor κB
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}