


Abstract
The γ-aminobutyric acid (GABA) receptor type B (GABABR) is constituted of at least two homologous proteins, GABABR1 and GABABR2. These proteins share sequence and structural similarity with metabotropic glutamate and Ca2+-sensing receptors, both of which are sensitive to Ca2+. Using rat brain membranes, we report here that the affinity of GABA and 3-aminopropylphosphinic acid for the GABABR receptor is decreased by a factor >10 in the absence of Ca2+. Such a large effect of Ca2+ is not observed with baclofen or the antagonists CGP64213 and CGP56999A. In contrast to baclofen, the potency of GABA in stimulating GTPγS binding in rat brain membranes is also decreased by a factor >10 upon Ca2+ removal. The potency for Ca2+ in regulating GABA affinity was 37 μM. In cells expressing GABABR1, the potency of GABA, but not of baclofen, in displacing bound 125I-CGP64213 was similarly decreased in the absence of Ca2+. To identify residues that are responsible for the Ca2+ effect, the pharmacological profile and the Ca2+ sensitivity of a series of GABABR1 mutants were examined. The mutation of Ser269 into Ala was found to decrease the affinity of GABA, but not of baclofen, and the GABA affinity was found not to be affected upon Ca2+ removal. Finally, the effect of Ca2+ on the GABAB receptor function is no longer observed in cells coexpressing this GABABR1-S269A mutant and the wild-type GABABR2. Taken together, these results show that Ser269, which is conserved in the GABABR1 protein fromCaenorhabditis elegans to mammals, is critical for the Ca2+-effect on the heteromeric GABAB receptor.
The neurotransmitter γ-aminobutyric acid (GABA) activates both ionotropic receptors (the GABAA and GABAC receptors) and G protein-coupled metabotropic receptors (the GABAB receptors). GABAA and GABAC receptors are permeable to Cl− ions and mediate, in most cases, fast inhibition of the postsynaptic neurons. GABAB receptors mostly couple to pertussis toxin-sensitive G proteins (Gi or Go) and accordingly inhibit the activity of adenylyl cyclase and voltage-gated Ca2+-channels or activate G protein-regulated inwardly rectifying K+-channels (Kir3) (for reviews, see Kerr and Ong, 1995; Bettler et al., 1998). When located on the postsynaptic element, the GABAB receptors inhibit neuronal excitability (Kerr and Ong, 1995; Bettler et al., 1998). GABAB receptors are also found on glutamatergic or GABA-ergic terminals, where they inhibit neurotransmitter release. GABAB receptors are widely expressed in the central nervous system, both in brain and spinal cord. Accordingly, these receptors play important physiological roles in pain transmission, absence epilepsy, and drug addiction (Kerr and Ong, 1995; Bettler et al., 1998).
Two GABAB receptor proteins have been identified: GABABR1 (Kaupmann et al., 1997) and GABABR2 (Jones et al., 1998; Kaupmann et al., 1998b; White et al., 1998; Kuner et al., 1999; Ng et al., 1999). Neither receptor was found to efficiently couple to G proteins when expressed alone (Kaupmann et al., 1997, 1998a,b; Martin et al., 1999). Indeed, GABAB receptors that efficiently activate pertussis-toxin-sensitive G proteins were only obtained in cells coexpressing GABABR1 and GABABR2 (Jones et al., 1998; Kaupmann et al., 1998b; White et al., 1998; Kuner et al., 1999; Ng et al., 1999). Moreover, these two receptors were found to associate at least by their carboxyl-terminal sequence (White et al., 1998; Kuner et al., 1999), further suggesting that GABAB receptors function in a heteromeric form. Both GABABR1 and R2 proteins share sequence similarity with family 3 G protein-coupled receptors (GPCRs) (Bockaert and Pin, 1999), which include metabotropic glutamate receptors (mGluRs), Ca2+-sensing receptors (CaSR) and putative pheromone and taste receptors. Like all family 3 GPCRs, GABAB receptors possess a large amino-terminal domain structurally related to some bacterial periplasmic proteins, where ligands bind (Galvez et al., 1999;Malitschek et al., 1999). This amino-terminal domain is attached to a heptahelical transmembrane domain that constitutes the G protein-activating domain, as demonstrated for mGluRs (Pin et al., 1994; Gomeza et al., 1996a).
Because GABAB receptors share sequence similarity with mGluRs and CaSRs, two types of receptors sensitive to Ca2+ ions (Brown and Hebert, 1997; Kubo et al., 1998; Saunders et al., 1998), we examined a possible regulation of GABAB receptors by Ca2+ions. We found that Ca2+ affects the affinity of various GABAB ligands differently and identified an amino acid residue in GABABR1 that is critical for this effect.
Experimental Procedures
Materials.
GABA was obtained from Sigma (L'Isle d'Abeau, France). 125I-CGP64213 was synthesized from ethyl-(1,1-diethoxyethyl)phosphonate as described elsewhere (Froestl et al., 1995) and labeled to a specific radioactivity of >2000 Ci/mmol (ANAWA AG, Wangen, Switzerland). All the other ligands were synthesized in-house (Froestl et al., 1995). Serum, culture media, and other solutions used for cell culture were from Life Technologies, Inc. (Cergy Pontoise, France).
Culture and Transfection of 293 Cells.
Human embryonic kidney (HEK) 293 cells were cultured in Dulbecco's modified Eagle's medium (Life Technologies, Cergy Pontoise, France) supplemented with 10% fetal calf serum and transfected by electroporation as described previously (Gomeza et al., 1996b). Electroporation was carried out in a total volume of 300 μl with 10 μg of carrier DNA, plasmid DNA containing the wild-type or mutated GABABR1 (1 μg) (Galvez et al., 1999), GABABR2 (1 μg) (Kaupmann et al., 1998a), or Gqi9 (1 μg) (Conklin et al., 1993) and 10 million cells.
Ligand Binding Assay.
Rat brain membranes were prepared as described previously (Olpe et al., 1990) and stored at −70°C. Membranes of HEK 293 cells were prepared as follows: 24 h after transfection, the cells were transferred into serum-free Dulbecco's modified Eagle's medium. Forty-eight hours after transfection, the cells were washed and homogenized in Tris-Krebs buffer [20 mM Tris-Cl, pH 7.4, 118 mM NaCl, 5.6 mM glucose, 1.2 mM KH2PO4, 1.2 mM MgSO4, 4.7 mM KCl, 1.8 mM CaCl2 (omitted for calcium-free experiments)] and centrifuged for 20 min at 40,000g. The pellet was resuspended in Tris-Krebs buffer and stored at −80°C. For ligand competition assays, thawed membranes (10 μg of protein) were incubated with 0.1 nM 125I-CGP64213 in the absence or presence of unlabeled ligands at different concentrations, as indicated on the figures. This concentration of125I-CGP64213 is about 10 times lower than the affinity of this radioligand on the native or GABABR1 receptor, such that the IC50 values measured for all unlabeled compounds are not significantly different from their affinity. The nonspecific binding was determined using 10 mM GABA or 100 nM CGP54626A. The incubation was terminated by filtration through GF/C Whatman glass fiber filters (Whatman International Ltd., Maidstone, England).
GTPγ35S Assay.
Rat brain synaptic membranes were prepared as described previously (Olpe et al., 1990) and stored in −70°C. To perform this assay on recombinant receptors, membranes were prepared from Chinese hamster ovary K1 cells (American Type Culture Collection, Manassas, VA) stably transfected with GABABR1b and GABABR2 cDNAs. Human GABABR1b and rat GABABR2 constructs [R1 in pcDNA3.1, Invitrogen (San Diego, CA); R2 in pC1-neo (Promega, Madison, WI) were cotransfected (1:1 ratio of plasmids) using the Superfect transfection system from Qiagen AG (Basel, Switzerland). Stably transfected cell clones were selected by double selection with 1 mg/ml geneticin and 250 μg/ml zeocin. Membranes were thawed using an ice-water bath and centrifuged at 4°C for 15 min at 20,000g. After addition of 1.2 ml of ice-cold water, the pellet was shaken, incubated on ice for 1 h, and centrifuged again. Afterward, the pellet was homogenized in guanosine 5′-O-(3-thiotriphosphate) (GTPγS) buffer (50 mM Tris-Cl, pH 7.7, 10 mM MgCl2, 0.2 mM EGTA, 100 mM NaCl) using a glass/Teflon homogenizer. Assay mixtures including membranes (20–30 μg) and test substances were prepared in Packard-Pico-Plates 96 (300-μl volume), supplemented with 30 μM GDP (Sigma) (10 μM in experiments conducted with recombinant receptors) and incubated for 30 min at room temperature. When experiments were conducted in the presence of Ca2+, CaCl2 was added at a concentration of 2.0 mM (final free Ca2+ concentration, 1.8 mM). The main incubation started by the addition of [35S]GTPγS (Amersham Paisley, UK) to a final concentration of 0.2 nM (0.3 nM in experiments conducted with recombinant receptors). Nonspecific binding was measured in the presence of 10 μM unlabeled GTPγS. After 40 min incubation at room temperature, the amount of bound [35S]GTPγS was assessed by standard procedures such as vacuum filtration through GF/B filters followed by liquid scintillation counting or scintillation proximity assay using 1.5 mg of wheat germ agglutinin-coated scintillation proximity assay beads (Amersham) per 250 μl of assay mixture.
Determination of Inositol Phosphate Accumulation.
Determination of Inositol phosphate (IP) accumulation in transfected cells was performed after labeling the cells overnight with myo-[3H]inositol (23.4 Ci/mol; NEN, Les Ullis, France). The stimulation was conducted for 30 min in a medium containing 10 mM LiCl and the indicated concentration of agonist. Results are expressed as the amount of IP produced over the radioactivity present in the membranes.
Data Analysis.
The curves were fitted with either GraphPAD Prism software (GraphPAD Software, San Diego, CA) or Kaleidagraph software (Abelbeck/Synergy Software, Reading, PA) using the equation y = {(ymax − ymin) / [1 + (×/EC50)nH]) + ymin}, where EC50 is the concentration of the compound necessary to obtain 50% of the maximal effect andn H is the Hill coefficient.
Results
Ca2+ Differently Regulates the Affinity of Several Compounds on Native GABAB Receptors.
The effect of Ca2+ removal on the affinity of several ligands interacting with the GABAB receptor was examined in rat brain membranes using 125I-CGP64213 as radioligand. This compound has previously been reported to selectively label GABABR1 proteins in the mammalian brain (Kaupmann et al., 1997). As shown in Table1, the potencies of cold CGP64213 and of CGP56999A, another antagonist, were only slightly affected upon removal of Ca2+ (1 mM EGTA added). A more pronounced effect of Ca2+ was, however, observed with the other antagonist CGP54626A. In contrast, the potency of the agonists GABA and APPA were decreased by a factor of more than 10 (Table 1). This effect is not a general effect for all GABABR agonists because the potency of baclofen was not affected by Ca2+ removal (Table 1). This indicates that Ca2+ is necessary for high-affinity binding of some, but not all, compounds interacting with native GABABR1. Because of the low concentration of125I-CGP64213 used (1/10 of itsK d value), the IC50values measured for unlabeled compounds are very near theirK i (affinity) values.
Effect of Ca2+ on the affinities of several GABABreceptor ligands on native GABAB receptors
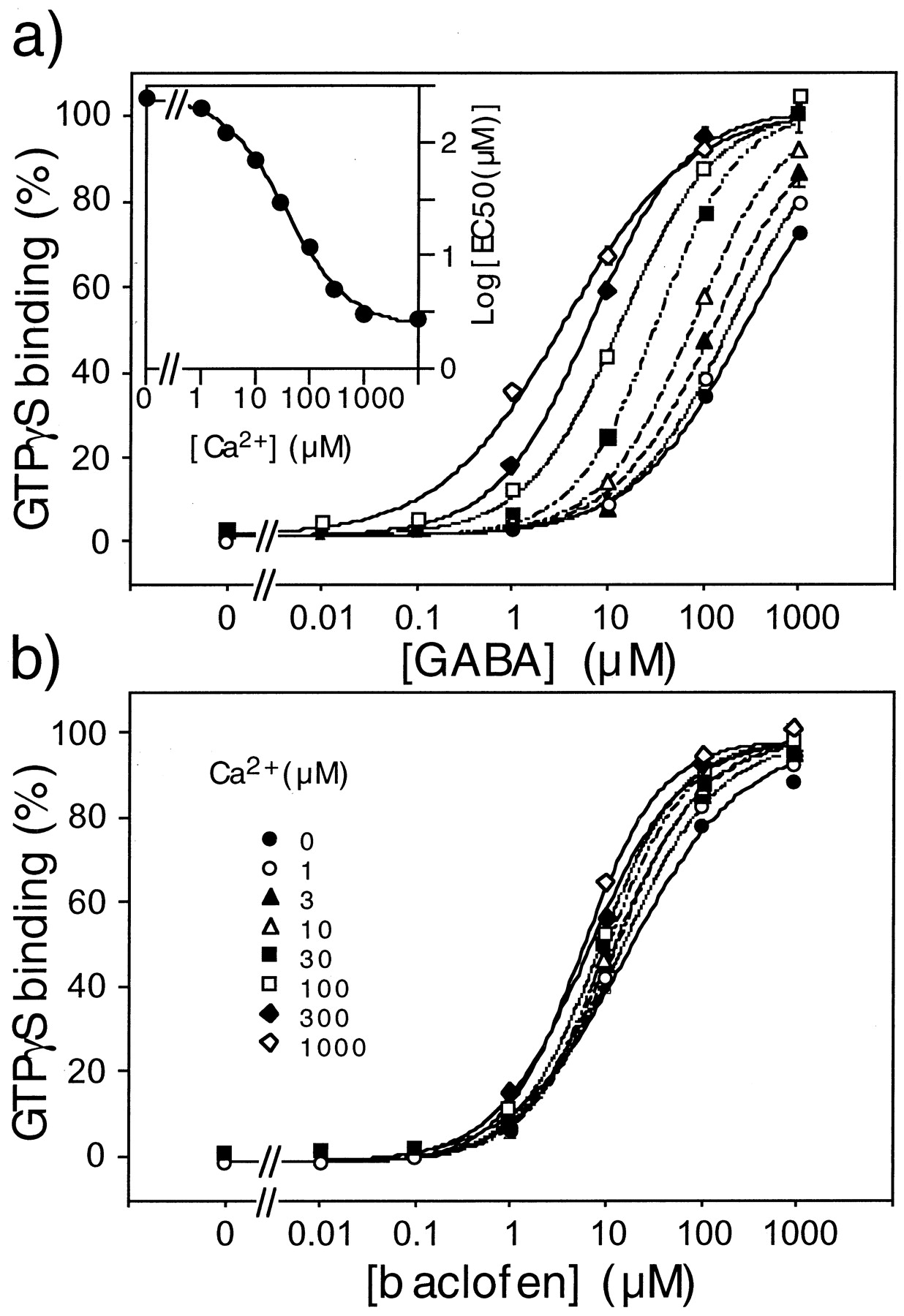
We also examined the effect of Ca2+ on the potency of GABA and baclofen to activate native GABAB receptors. In the presence of 1 mM Ca2+, GABA, APPA, and baclofen stimulated GTPγS binding in a dose-dependent manner (Table2). In the absence of Ca2+, the EC50 values for GABA and APPA were increased by a factor 80 whereas that of baclofen was only slightly affected (Fig. 1). When the Log(EC50) values for GABA determined at various Ca2+ concentrations were plotted against the Ca2+ concentration, an EC50 value of 37 ± 5 μM was determined for Ca2+ (Fig. 1a, inset).
Effect of Ca2+ on the potencies of several GABABreceptor agonists in stimulating [35S]GTPγS binding on native and recombinant GABAB receptors

Effect of Ca2+ on GABA (a) and baclofen (b) stimulated GTPγS binding on rat brain membranes. The binding of [35S]GTPγS on rat brain membranes was measured in the presence of the indicated concentration of GABA (a) or baclofen (b), and in the presence of the indicated concentration of Ca2+: 0 μM (●), 1 μM (○) 3 μM (▴), 10 μM (▵), 30 μM (▪) 100 μM (▪) 300 μM (♦), 1000 μM (⋄). Data are means ± S.E.M. of a triplicate determination from a typical experiment. Inset in a: plot of the EC50 value for GABA as a function of Ca2+ concentration.
Effect of Ca2+ on the Recombinant GABABReceptors.
Because 125I-CGP64213 has been reported to bind on the GABABR1 subunit but not on the GABABR2 subunit (Kaupmann et al., 1998a), we first examined the effect of Ca2+ on the recombinant GABABR1a subunit expressed alone in HEK 293 cells. As shown in Table 1, the affinities of CGP64213, CGP56999A, and baclofen were not affected by removal of Ca2+, but those of GABA and APPA were largely decreased (Fig. 2 and Table 1). Similar data were obtained with the GABABR1b splice variant, which lacks the 120 amino-terminal residues that correspond to the sushi domains of GABABR1a (data not shown) (Kaupmann et al., 1997; Hawrot et al., 1998).

Inhibition curves of 125I-CGP64213 binding on membranes prepared from cells expressing GABABR1a by GABA and baclofen in the presence (a) and absence (b) of Ca2+. The binding of 0.1 nM125I-CGP64213 on HEK 293 cell membranes (10 μg) was measured in the presence of the indicated concentration of GABA (○) or baclofen (●), and in the presence (a) or absence (b, 1 mM EGTA added) of added Ca2+ (1 mM). Data are means ± S.E.M. of triplicate determinations from a single typical experiment.
As observed with the native receptors, Ca2+ was also required for the high potency of GABA and APPA to stimulate [35S]GTPγS binding on membranes prepared from cells stably expressing both GABABR1 and GABABR2 subunits (Table 2). On these membranes, the effect of baclofen was found to be unaffected by Ca2+ removal (Table 2). The shift in GABA and APPA potencies observed upon removal of Ca2+ was, however, found to be lower on the recombinant receptor than on the native receptors (Table 2).
Effect of Point Mutations in GABABR1 on the Ca2+-Effect.
Because Ca2+affects both the potency measured in a functional assay and the affinity of GABA and APPA, it is likely that Ca2+acts directly on the ligand-binding suppress domain of the GABABR1 protein. We therefore first examined whether the effect of Ca2+ could still be observed on a truncated GABABR1 receptor protein that corresponds to the LBP-like domain attached to the plasma membrane by only one transmembrane segment. We previously reported that such a truncated receptor still binds 125I-CGP64213 (Malitschek et al., 1999). On such a truncated protein, the affinity of GABA, but not of baclofen, was also decreased in the absence of Ca2+ (data not shown), indicating that Ca2+ interacts on the LBP-like domain of the GABABR1 protein.
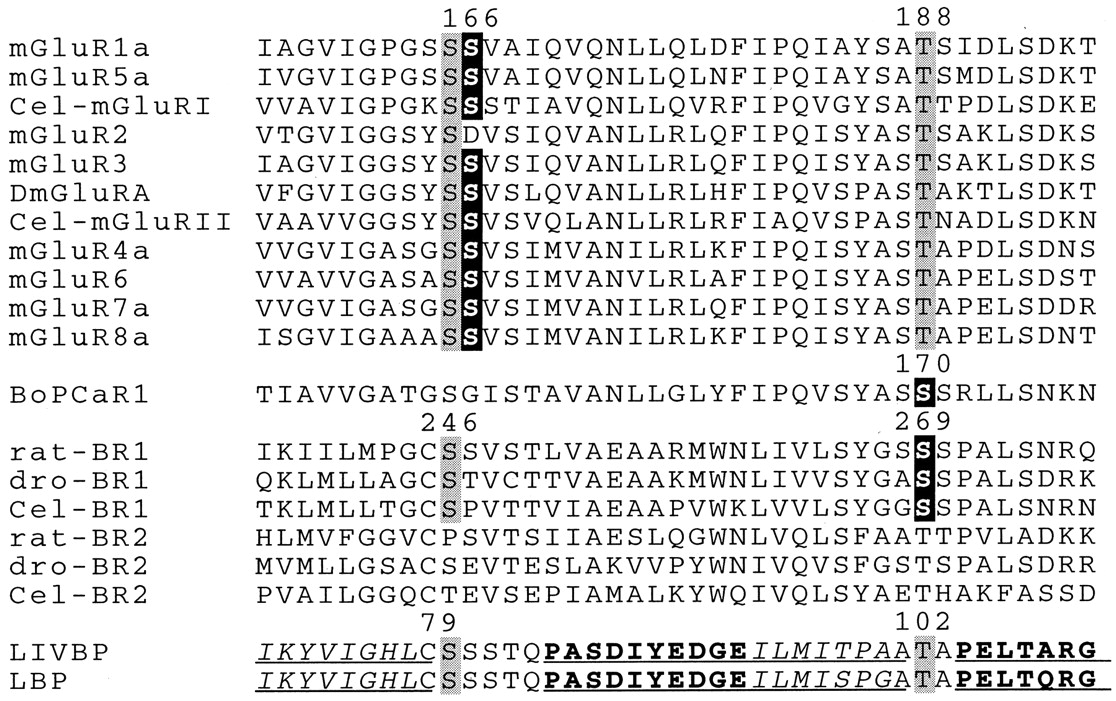
Because of the specific effect of Ca2+ on the affinity of some but not all GABAB ligands, we speculated that the amino acid residues responsible for the Ca2+ effect are near the GABA binding site. In accordance with this idea, a single residue (Ser166) in mGluR1, located near a serine residue involved in glutamate binding (Ser165), plays a critical role in the Ca2+ effect on this receptor subtype (Kubo et al., 1998). These two serine residues are conserved in GABABR1a (Ser247 and Ser246, respectively) (Fig.3), and using molecular modeling and mutagenesis, we showed that Ser246 is probably directly involved in the binding of GABA (Galvez et al., 1999). One would therefore predict that Ser247 could be responsible for the Ca2+ effect in GABABR1a. In addition to Ser247, at least two other serine residues (Ser268 and Ser269) are also near Ser246 in our three-dimensional model of the GABABR1a binding site. Interestingly, Ser269 of GABABR1a aligned with a serine residue of the CaSR that plays an important role for its activation by Ca2+ (Bräuner-Osborne et al., 1999) (see Fig. 3).

Alignment of the mGlu, Ca-sensing and GABAB receptor sequences with those of the periplasmic binding proteins LIVBP and LBP, in one of their regions responsible for ligand binding. Residues highlighted in gray have been shown (for LIVBP and LBP) or proposed to directly interact with the ligand (Glu in mGluRs, GABA in GABABR1, and leucine in LIVBP). Residues highlighted in black are responsible for the effect of Ca2+in mGluRs and CaSR, and that we identified as critical for the effect of Ca2+ on GABABR1. The numbers on top of the sequences correspond to the position of the residues in the rat mGluR1, bovine CaSR, rat GABABR1a (all including the signal peptide sequence) and LIVBP (from top to bottom). The known secondary structural elements of LIVBP and LBP are underlined (β strands in italic, and α helices in bold). The sequences of the D. melanogaster (dro) GABABR1 and GABABR2 were obtained from cosmids DS00929 and DS02649, respectively. TheC. elegans (Cel) sequences Cel-BR1 and Cel-BR2 were obtained from cosmids Y41G9 and ZK180, respectively. The C. elegans sequences Cel-mGluRI and Cel-mGluRII were obtained from the cosmids F45H11 and ZC506, respectively. The alignment was generated using the sequence of the LBP-like domain of the receptors with the default parameters of the Clustal-W program.
Therefore, the effect of the mutation of Ser247, Ser268, and Ser269 into Ala on the potency of several GABAB ligands to displace 125I-CGP64213 specific binding was examined. The potency of CGP64213 was not significantly affected in any of these mutated receptors [IC50 = 1.44 ± 0.33, 0.94 ± 0.15, and 4.48 ± 0.84 nM (n = 3) on GABABR1a-S247A, -S268A, and -S269A mutants, respectively, compared with 1.53 ± 0.25 nM for the wild-type receptor]. Moreover, the potency of CGP64213 was affected by a factor of ∼2 upon removal of Ca2+(IC50 = 3.25 ± 0.40, 1.98 ± 0.17, and 7.04 ± 2.11 nM (n = 3) on GABABR1a-S247A, -S268A, and -S269A mutants, respectively).
If one of the above-mentioned residues is involved in the Ca2+-sensitivity of GABABR1a, its mutation into an alanine is expected to have an effect similar to that resulting from Ca2+ removal: a large decrease in the affinity for GABA but not for baclofen. Indeed, among the three mutants, only GABABR1a-S269A displayed these features, as reported previously (Galvez et al., 1999) (Table3). Moreover, Ca2+removal from the incubation buffer resulted in a decreased affinity for GABA in GABABR1a-S247A and GABABR1a-S268A mutants but not in the GABABR1a-S269A mutant (Fig.4 and Table 3), further indicating that Ser269 plays a critical role in the Ca2+ effect.
Effect of Ca2+ on the affinities and potencies of GABA and baclofen on wild-type and mutant GABABR1a receptors

Ca2+-sensitivity of GABA binding on S247A (a), S268A (b), and S269A (c) GABABR1a mutants. The binding of 0.1 nM 125I-CGP64213 on membranes prepared from HEK 293 cells expressing the indicated receptor was measured in the presence of the indicated concentration of GABA (○) or baclofen (●), and in the presence (left) or absence (plus 1 mM EGTA) of added Ca2+(1 mM). Data are means ± S.E.M. of a triplicate determination from a typical experiment. For each mutant, the data with GABA and baclofen with and without Ca2+ were obtained in the same experiment.
Ser269 of the GABABR1 Receptor Subunit Plays a Critical Role for the Ca2+-Effect on the Heteromeric GABAB Receptor.
The effect of the mutation of Ser269 in GABABR1a was examined on the functional activity of the heteromeric GABABR1/GABABR2 receptor. To that aim, GABABR1a and GABABR2 were coexpressed in HEK 293 cells together with the chimeric G protein α-subunit Gqi9 [Gαq in which nine residues of the carboxyl-terminus have been replaced by those of Gαi (Conklin et al., 1993)]. Under these conditions, the GABAB receptor was found to stimulate phospholipase C (PLC), and display pharmacological properties similar to those measured using other functional assays (Franek et al., 1999). Using this assay, and as observed with other functional readouts, GABA and baclofen display a similar potency in the presence of 1 mM Ca2+ (Fig. 5 and Table 3). In the absence of extracellular Ca2+, GABA was 10 times less potent than baclofen (Fig. 5a and Table 3), as observed with the native receptor or the recombinant heteromeric receptor using the GTPγS binding assay (Table 2).

Effect of Ca2+ on GABA and baclofen induced activation of wild-type or GABABR1a-S269A + GABABR2 receptors. The IP formation induced by various concentrations of GABA (○) or baclofen (●) was measured on HEK 293 cells expressing the chimeric G protein Gqi9, GABABR2, and either the wild-type GABABR1a (a) or the GABABR1a-S269A mutant (b). The experiment was conducted either in the presence (left) or absence (plus 1 mM EGTA) of added Ca2+ (1 mM). Data are means ± S.E.M. of a triplicate determination from a typical experiment. For each mutant, the data obtained with GABA and baclofen, with and without Ca2+ were obtained in the same experiment.
However, using the PLC assay, we found that the potency of baclofen was also decreased in the absence of Ca2+, although to a lower extent than that of GABA (Table 3). Because this effect was observed neither on the potency of baclofen to displace125I-CGP64213 binding nor on its potency to stimulate [35S]GTPγS binding (see above), it probably resulted from a Ca2+ requirement in a step downstream of the G protein activation by the GABAB receptor. Indeed, Ca2+ is known to have a positive effect on PLC activity (Cockcroft and Thomas, 1992) such that the removal of Ca2+ from the external milieu can affect the coupling efficacy of a receptor to PLC, leading to a decrease in both potency and efficacy of agonists. In agreement with this proposal, the maximal effect not only of baclofen but also of GABA or APPA was decreased by 2- to 3-fold (Fig. 5A), indicating it is a general feature of the PLC cascade activation upon stimulation of the receptors by any agonist. Moreover, the effect of Ca2+ on the potency of baclofen in stimulating IP formation occurred in a concentration range (30–100 μM) different from that affecting the potency of GABA (1–30 μM) in this assay. According to this additional effect of Ca2+ on the PLC activation, the effect of Ca2+ on the GABA potency on wild-type and mutated receptors was always compared with that of baclofen.
GABA and baclofen still activated in a dose-dependent manner the heteromeric GABAB receptors, constituted of the wild-type GABABR2 and either the GABABR1a-S247A or GABABR1a-S268A mutant. These two agonists displayed a similar affinity on these two mutant receptors in the presence of Ca2+, whereas GABA became much less potent than baclofen in the absence of Ca2+(Table 3). These data are very similar to those obtained with the wild-type receptor, further indicating that these two serine residues are not involved in the Ca2+-sensitivity of the GABA activation of the heteromeric receptor.
In cells expressing the GABABR1a-S269A mutant and GABABR2, we noticed a decrease by about a factor 10 of the GABA potency but no change in the EC50value for baclofen compared with the wild-type receptor (Fig. 5 and Table 3). Interestingly, this difference in the GABA potency mirrors the change in potency of GABA observed on the wild-type receptor upon Ca2+ removal. In addition, the relative potency of GABA over that of baclofen on cells expressing GABABR1a-S269A and GABABR2 remained unchanged in the absence of external Ca2+ (Fig. 5b and Table 3). This shows that the GABABR1a-S269A/GABABR2 heteromer is no longer sensitive to Ca2+.
In brain membranes, we noticed that the removal of Ca2+ largely decreased not only the affinity of GABA, but also that of APPA. If Ser269 is playing a critical role for the Ca2+ effect on the GABAB receptor, not only the affinity of GABA, but also that of APPA should be affected by this mutation. As shown in Table 4, this is exactly what we observed. Moreover, this mutation largely decreased the effect of Ca2+ on the affinity of APPA (Table 4). Surprisingly, whereas the affinity of CGP54626A and CGP56999A on either wild-type or recombinant receptors is slightly decreased upon Ca2+ removal (Table 1), the mutation of Ser269 into alanine resulted in a large decrease in the affinity of CGP54626A but not of CGP56999A (Table 4). This revealed that the effect of the mutation of Ser269 on the affinity of CGP54626A cannot be simply explained by the suppression of the action of Ca2+ on the GABABR1 subunit.
Effect of Ca2+ on the affinities of several compounds on the GABABR1a S269A mutant compared with the wild-type receptor
Discussion
The present data show that Ca2+ is necessary for the high affinity of GABA and APPA on native and recombinant GABAB receptors. Surprisingly, the action of baclofen on this receptor does not require Ca2+ions. Such a specific decrease in GABA and APPA affinities on wild-type receptor observed upon Ca2+ removal was mimicked by mutating the Ser269 of GABABR1a into Ala. Moreover, Ca2+ removal has no further effect on the GABA and APPA affinities on this mutant receptor. These data demonstrate that Ser269 in the GABABR1 subunit is responsible for the Ca2+ effect on the heteromeric GABAB receptor. In agreement with an important role for Ser269 in GABABR1, this specific residue is conserved in both the Caenorhabditis elegans and Drosophila melanogasterGABABR1 receptors (Fig. 3). In contrast, Ser247 and Ser268, which we show are not important for the Ca2+-effect on GABABR1 are not conserved in the C. elegans and D. melanogaster sequences (Fig. 3).
Comparison with CaSR and mGluRs.
Within the different members of the family 3 GPCRs, Ca2+ has been shown to regulate the activity of the CaSR and of some mGluRs. In the case of the CaSR, Ca2+ seems to be sufficient to activate the receptor (Brown et al., 1993; Hammerland et al., 1999). In the case of mGluR1, mGluR3, and mGluR5, Kubo et al. proposed that Ca2+ also directly activates these receptors (Kubo et al., 1998). However, other authors did not observe such a direct activation of mGluR1 by Ca2+; rather, they found that Ca2+ increases the potency of glutamate to activate the receptor (Saunders et al., 1998). These later observation are not in contrast to those reported by Kubo et al., because it is possible that in Kubo's experiments, the extracellular glutamate concentration was too low to activate the receptor in the absence of Ca2+, but became effective upon addition of Ca2+. Ca2+seems, therefore, to exert a similar action on some mGluRs and GABAB receptors acting as an allosteric regulator and increasing the potency of some agonists. However, the apparent affinity of Ca2+ on mGluRs and GABAB receptors is quite different, being in the millimolar range for mGluRs (Kubo et al., 1998; Saunders et al., 1998) and around 37 μM for GABAB receptors, suggesting that Ca2+ may not interact with these two receptor types in the same way.
Possible Mode of Action of Ca2+ on GABABR1.
Ca2+ has also been shown to act on the extracellular leucine/isoleucine/valine binding protein (LIVBP)-like domain of the CaSR (Hammerland et al., 1999). Recently, Ser170 has been reported to play a critical role for the action of Ca2+ (Bräuner-Osborne et al., 1999). Interestingly, this residue aligns with Ser269 of GABABR1a (Fig. 3). However, the difference in potencies of Ca2+ on these two receptors (37 μM on GABABR and 3 mM on CaSR), plus the observation that Ca2+ is sufficient to activate CaSR but not GABABR, suggest that Ca2+does not act similarly on these two receptors.
In mGluRs, the absence of effect of Ca2+ on mGluR2 allows the identification of a single residue conserved in mGluR1, mGluR3, and mGluR5, and responsible for the Ca2+-effect on these receptors (Kubo et al., 1998). This residue is Ser166 in mGluR1, which is replaced by an Asp in mGluR2 (see Fig. 3). Ser166 is very close to the glutamate binding site, because the α-carboxylic and α-amino groups of glutamate have been proposed to interact with Ser165 and Thr188, respectively (O'Hara et al., 1993) (Fig. 3). In GABABR1a, Ser246 aligns with Ser165 of mGluR1 and has been reported to play a critical role for the binding of GABA and other GABABligands (Galvez et al., 1999). These two residues align with Ser79 of LIVBP (Fig. 3), which forms a hydrogen bound with the α-carboxylic group of leucine in the bacterial protein (Sack et al., 1989), suggesting that the glutamate and GABA binding sites in mGluRs and GABABR1a may be similar to that of the leucine binding site in LIVBP. Although Ser166 of mGluR1 is conserved in both mammalian GABABR1a (Ser247) and GABABR2 (Ser137), this residue is not involved in the Ca2+ effect on the GABAB receptor. Indeed, as summarized above, our data indicate that Ser269 of GABABR1a is responsible for the Ca2+ effect. Ser269 of GABABR1a aligns with Thr188 of mGluR1 and Thr102 of LIVBP (Fig. 3). In mGluR1, Thr188 has been shown to interact with glutamate (O'Hara et al., 1993), and in LIVBP, Thr102 forms a hydrogen bond with both the α-amino and α-carboxylic groups of leucine (Sack et al., 1989). Because GABA lacks the α-amino group common to both leucine and glutamate, two possible modes of action of Ca2+ on the GABAB receptor can be proposed. Ca2+ could compensate the lack of the α-amino group of GABA, interacting with Ser269 and allowing a better positioning of the carboxylic group of GABA for its proper interaction with Ser246. Alternatively, Ca2+ may not interact directly with Ser269 but may affect the position of its OH group, allowing the formation of an additional hydrogen bond with the carboxylic group of GABA. Further experiments are required to clarify the mode of action of Ca2+ on the GABAB receptor.
The observation that baclofen does not require Ca2+ for high-affinity binding on GABABR1 may be explained in two ways. One possibility is that, because of the additional chlorophenyl group of this molecule, baclofen does not interact the same way as GABA in GABABR1a. Alternatively, the presence of the chlorophenyl group may not allow the binding of Ca2+ on the receptor.
Possible Physiopathological Relevance of the Ca2+-Effect on GABAB Receptors.
The potency of Ca2+ on both CaSR and mGluRs is in the millimolar range (Brown and Hebert, 1997; Kubo et al., 1998); therefore, it is in the range of the Ca2+concentrations found in blood and in cerebrospinal fluid. Accordingly, the Ca2+-sensing function of both CaSR and mGluRs may play important physiological roles. In the case of the CaSR, this has been pointed out by identifying many point mutations in the CaSR gene that affect the potency of Ca2+ to activate this receptor, and that are associated with genetic diseases affecting the regulation of calcemia (Pollak et al., 1993; Pollak et al., 1994;Brown and Hebert, 1997). In the case of the GABABreceptor, we found that the EC50 value for Ca2+ is 37 μM, such that a significant shift of the GABA dose-response curve can only be detected when the extracellular Ca2+ concentration was lowered to 100 μM. In the brain, such low extracellular Ca2+ concentrations can be reached only under particular conditions, such as activation of glutamate receptors, especially those of the NMDA type (Pumain et al., 1983; Pumain and Heinemann, 1985; Pumain et al., 1987), or after epileptic seizure (Pumain et al., 1983; Heinemann et al., 1986; Armand et al., 1995). If this occurs in the vicinity of the GABABreceptors, it will result in a decreased activity of GABAB receptors, thereby preventing this receptor from being activated by GABA. Because GABABreceptors regulate both glutamatergic and GABA-ergic synaptic transmission, it is difficult to predict whether such an effect will facilitate or inhibit the development of the seizure.
In conclusion, we show here that Ca2+ ions are required for the high-affinity binding of some but not all GABAB ligands, and we show that Ser269 of the GABABR1 subunit is involved in this Ca2+-effect. Because of the high potency of Ca2+ for the GABABreceptor, this effect is unlikely to have an important regulatory role. However, one should take into account the presence of Ca2+ in the GABABR1 binding site for the understanding of the activation process of GABAB receptors.
Acknowledgments
We thank Dr. Joëll Bockaert for constant support during this work and Audrey Boncompain and Karin Hofstetter for technical help.
Footnotes
- Received July 30, 1999.
- Accepted October 20, 1999.
-
Send reprint requests to: Dr. Thierry Galvez, CCIPE-Centre National de la Recherche Scientifique-UPR9023, Laboratoire des Mécanismes Moléculaires des Communications Cellulaires, 141 rue de la Cardonille, F-34094 Montpellier Cedex 5, France. E-mail:galvez{at}ccipe.montp.inserm.fr
-
This work was supported by grants from the “Action Incitative Physique et Chimie du vivant” (PCV97–115), the European Community Biomed2 (BMH4-CT96–0228) and Biotech2 (BIO4-CT96–0049) programs, the Fondation pour la Recherche Médicale, Novartis Pharma (Basel, Switzerland) and the association Retina France (all to J.-P.P.)
Abbreviations
- GABA
- γ-aminobutyric acid
- GPCR
- G protein-coupled receptor
- mGluR
- metabotropic glutamate receptor
- CaSR
- Ca2+-sensing receptor
- HEK
- human embryonic kidney
- GTPγS
- guanosine 5′-O-(3-thiotriphosphate)
- IP
- inositol phosphates
- APPA
- 3-aminopropylphosphinic acid
- LBP
- leucine binding protein
- PLC
- phospholipase C
- LIVBP
- leucine/isoleucine/valine binding protein
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}