


Abstract
A molecular model of the human melanin-concentrating hormone (MCH) peptide was constructed and docked into a helical, bacteriorhodopsin-based model of the recently identified human MCH receptor. From this hormone-receptor complex, potential sites of agonist-receptor interaction were identified, and site-directed mutagenesis was used to substitute residues predicted to reside within the receptor binding pocket. Substitution of Asp123(3.32) in the third transmembrane domain of the receptor resulted in a loss of detectable 125I-MCH binding and of MCH-stimulated Ca2+ flux; cell surface expression of the mutant receptor was not affected. Arg11 and Arg14 of the MCH ligand were identified as potential sites of interaction with Asp123(3.32). [Ala14]-MCH was equipotent to native MCH in its ability to bind to and activate the wild-type MCH receptor, whereas [Ala11]-MCH displayed a 3000-fold reduction in binding affinity and a complete loss of measurable functional activity. Furthermore, [Lys11]-MCH and [d-Arg11]-MCH displayed reduced affinity for the receptor. [Lys11]-MCH was observed to be a partial agonist, eliciting approximately 67% of the native peptide's activity in a Ca2+ flux assay, and [d-Arg11]-MCH was determined to be a functional antagonist with a K b valve of 15.8 μM. These data provide evidence that a basic moiety with specific stereochemical requirements at this site is needed for receptor activation. We conclude that both Asp123(3.32) in the MCH receptor and Arg11 in the MCH peptide are required for the formation of the MCH peptide/receptor complex and propose that they form a direct interaction that is critical for receptor function.
Melanin-concentrating hormone (MCH; NH2-Asp-Phe- Asp-Met-Leu-Arg-Cys-Met-Leu-Gly-Arg-Val-Tyr-Arg-Pro- Cys-Trp-Gln-Val-COOH) is a cyclic hypothalamic neuropeptide that was first characterized in the chum salmon pituitary as a hormone responsible for color changes in response to environmental backgrounds (Kawauchi et al., 1983). In mammals, MCH appears to have evolved into an important regulatory hormone in feeding behavior and is not found in normal skin melanophores (Knigge et al., 1996). Evidence supporting a role for MCH in feeding includes the observation of increased feeding behavior in rats after direct i.c.v. administration of MCH (Rossi et al., 1997), the up-regulation of MCH mRNA during fasting and in obeseob/ob (leptin deficient) mice (Qu et al., 1996), a functional antagonistic effect of MCH on the action of melanocortin in the hypothalamus (Ludwig et al., 1998), and, most recently, the characterization of a MCH knockout mouse displaying reduced body weight due to hypophagia and an increase in metabolic rate (Shimada et al., 1998). There has been considerable interest in the potential for the development of antiobesity agents that inhibit these processes and, thus, in the identification of the receptor for MCH.
Efforts to identify the ligands for orphan G protein-coupled receptors have recently led to the discovery of the receptor for MCH. Concurrently, two different groups reported the identification of the human MCH receptor (Chambers et al., 1999; Saito et al., 1999) as the orphan G protein-coupled receptor SLC-1 (Kolakowski et al., 1996). The receptor was reported to bind MCH with nanomolar affinity and to be localized to specific brain regions implicated in feeding behavior, such as the ventromedial and dorsomedial nuclei of the hypothalamus, the hippocampus, the olfactory tubercle, and the nucleus accumbens. Interestingly, the MCH receptor has also been observed in areas associated with the dopaminergic system, such as the substantia nigra, amygdala, and ventral tegmental area. Since these initial reports, three other groups have published the identification by similar methods of MCH as the ligand for the rat homolog of SLC-1 (Bachner et al., 1999; Lembo et al., 1999; Shimomura et al., 1999).
The discovery of the MCH receptor permits detailed characterization and localization of ligand binding domains on the receptor protein. This is an important step in understanding the relationship between receptor structure and function and provides insights into the mechanism of receptor activation. Here we have undertaken the design of a molecular model of the MCH receptor based on bacteriorhodopsin and docked the cyclic nonadecapeptide MCH ligand into the putative transmembrane binding domain. From this model we have identified several candidate residues as potential sites of interaction with the ligand. Biochemical analysis of the receptor and the ligand by site-directed mutagenesis has identified Asp123(3.32) in the third transmembrane domain of the receptor as a critical residue involved in ligand binding and receptor activation. Additionally, Arg11 in the MCH peptide was determined to be important for receptor binding. Using this information, we have designed and characterized a novel competitive antagonist of the MCH receptor.
Materials and Methods
Molecular Modeling of the MCH Peptide/Receptor Complex.
A molecular model of the human MCH peptide ligand was constructed from the postequilibrium geometry observed in a molecular dynamics study byPaul and et al. (1990). The initial model was constructed with Sybyl/Biopolymer (Tripos Associates, St. Louis, MO) and refined with InsightII/Discover (MSI, San Diego, CA). Three adjustments were made: correction of the N-terminal extension overlap, side-chain adjustment, especially with respect to the Tyr13-OH:Cys7-O hydrogen bond, and a disulfide bond formation. This model was then refined by molecular mechanics minimization for 500 steps with Discover.
We then built a template-based homology model of the helical regions of the MCH receptor, using the atomic resolution coordinates of bacteriorhodopsin (Henderson et al., 1990) with Look (MAG, Palo Alto, CA), into which we docked the above MCH peptide ligand structure. This approach has been shown to be useful for the identification of ligand binding sites for many G protein-coupled receptors (Schwartz, 1994). Bacteriorhodopsin was used instead of rhodopsin because of the higher resolution structure available. This initial model was refined by 500 cycles of molecular mechanics minimization with InsightII/Discover after manual adjustment of side-chain geometry to reduce steric overlap. Because a portion of the MCH peptide extended beyond the helical bundle into the vicinity of the extracellular loops, we then constructed the extracellular loops of the receptor about the MCH docked helical receptor model with InsightII/Homology, and after manual adjustments, the model was refined by 500 steps of molecular mechanics minimization with InsightII/Discover. This refinement did not significantly alter the backbone geometry or relative positions of the peptide or receptor helices in the model.
Construction of HA-Tagged Wild-Type and Mutant MCH Receptors.
The wild-type human MCH receptor full-length cDNA was obtained from Human Genome Sciences (clone ID no. HSXAI35) and transferred into pcDNA3.1 vector (Invitrogen, Carlsbad, CA) by polymerase chain reaction, adding a Kozak sequence (Kozak, 1986) before the starting Met, but with no 5′ or 3′ untranslated regions. The MCH receptor amino terminus was epitope tagged by inserting the hemagglutinin (HA) protein epitope (residues 98–106 = YPYDVPDYA), using a polymerase chain reaction-based site-directed mutagenesis system (Excite; Stratagene, La Jolla, CA). Subsequent mutagenesis was performed on the HA-tagged MCH receptor, using site-directed mutagenesis strategies in which 33-bp primers were used to introduce the desired mutation (QuikChange; Stratagene). The resultant plasmid cDNAs were used to transform MAX Efficiency DH5α competent cells (Life Technologies, Grand Island, NY), and the cDNAs were purified using an anion exchange procedure (Qiagen, Valencia, CA). All nucleotide sequences of the mutated cDNAs were verified using the dRhodamine Terminator Cycle Sequencing Reaction system (PE Biosystems, Foster City, CA) and analyzed on an ABI PRISM 377 automated DNA sequencer (PE Biosystems).
Transfection of HA-Tagged Wild-Type and Mutant MCH Receptors.
MCH receptor cDNAs were transiently expressed in COS-7 cells using the LipofectAMINE reagent system (Life Technologies) for initial studies. For selected receptors, stable transfectants of Chinese hamster ovary (CHO) cells were obtained using 10 μg plasmid cDNA and the LipofectAMINE reagent system (Life Technologies), with subsequent selection in 500 μg/ml G418 (Geneticin; Life Technologies) and clonal selection using fluorescence analysis cell sorting (FACS) with anti-HA-fluorescein mouse monoclonal antibody, clone 12CA5 (Roche Molecular Biochemicals, Indianapolis, IN). All stably transfected CHO cells were maintained in F-12 nutrient mixture (Ham) media supplemented with 10% fetal bovine serum, 100 U/ml penicillin, 100 μg/ml streptomycin, and 500 μg/ml G418 (complete F-12 media).
Peptide Synthesis.
MCH peptide and its analogs were assembled from solid support, using standard 9-fluorenylmethoxycarbonyl chemistry on a model ABI 431A peptide synthesizer (PE Biosystems) or a model MOS496 multiple synthesizer (Advanced ChemTech, Louisville, KY). The peptides in acyclic, reduced form were cleaved and deprotected from the solid support by concentrated trifluoroacetic acid in the presence of proper scavengers (90% trifluoroacetic acid:2.5% H2O, 2.5% triethylsilane, 2.5% thioanisole, 2.5% ethanedithiol). The reduced peptides were purified by HPLC, and the molecular masses were confirmed by electrospray mass spectrometry. The reduced peptides were then oxidized either by air overnight at diluted concentrations (0.5–1 mg/ml) in ammonium bicarbonate buffer, pH 8.5, or by a disulfide-exchanging agent (e.g., 2,2′bispyridyl disulfide) (Maruyama et al., 1999). The disulfide-cross-linked, monomeric peptides were purified using HPLC, and their molecular masses were confirmed by electrospray mass spectrometry.
Saturation and Competition Binding Assays.
Wild-type or mutant MCH receptor transfected CHO cell membranes were made by harvesting stably transfected cells with a nonenzymatic cell dissociation buffer (Life Technologies), which were pelleted by centrifugation at 500g for 10 min, resuspended in lysis buffer [5 mM HEPES, pH 7.4, supplemented with protease inhibitor cocktail (Complete, EDTA-free; Roche Molecular Biochemicals)], and incubated on ice for 30 min. Membranes were collected by centrifugation at 11,000g for 30 min, resuspended in lysis buffer, and drawn through a 23-gauge needle five times. Aliquots were flash-frozen and stored at −80°C until use.
CHO cell membranes were thawed and diluted in 25 mM HEPES (pH 7.4), 10 mM MgCl2, 5 mM MnCl, and 10 mM NaCl supplemented with 0.1% BSA and protease inhibitor cocktail (Complete, EDTA-free, Roche Molecular Biochemicals) for each binding assay. For saturation binding, increasing concentrations of 125I-MCH (NEN Life Science Products, Boston, MA) were added to this resuspension and incubated for 30 min at room temperature with constant agitation in the presence or absence of 1 μM nonradiolabeled MCH. For competition binding, membranes were incubated with 250 pM125I-MCH and increasing concentrations of nonradiolabeled MCH or MCH peptide analog and incubated for 30 min at room temperature with constant agitation. After incubation, the membranes were filtered through a 10 mg/ml BSA-treated Whatman GF/B filter paper, washed three times with 4°C 25 mM HEPES (pH 7.4), 10 mM MgCl2, 5 mM MnCl, and 10 mM NaCl, with a Brandel Harvester apparatus (Gaithersburg, MD), and the radioactivity was quantified by γ-spectrometry. Data shown are the average of triplicate determinations and were repeated at least three independent times unless otherwise noted. The values reported were calculated from curve fitting the data with the program GraphPad Prism (GraphPad Software, San Diego, CA).
Competition binding assays of mutant MCH receptors transiently expressed in COS-7 cells were performed using similar methods.
Inositol Phosphate Assay.
Stably transfected CHO cells were plated in six-well culture dishes, grown to 80% confluency, and then equilibrated in complete F-12 media with 1 μCi/ml myo-[3H]inositol (NEN Life Science Products) overnight. Each well was washed with 37°C PBS (Life Technologies) and stimulated with agonist peptide in PBS containing 20 mM LiCl and 1 mM CaCl2 for 45 min at 37°C. Cell stimulation was terminated by aspiration of the peptide-containing buffer solution followed by the addition of 0.4 M perchloric acid (1.0 ml) to each well. The cell lysate was then neutralized by a 3:1 dilution with a 0.72 M KOH:0.6 M KHCO3 solution. Precipitate was removed by centrifugation (5000g, 15 min), and the supernatant was subjected to Dowex anion exchange chromatography, using AG 1-X8 resin, formate form (Bio-Rad, Hercules, CA) to separate inositol phosphates from free inositol. After copious washes with room temperature water (2 × 10 ml), inositol phosphates were eluted by the addition of a 0.1 M formic acid, 1 M NH4COOH solution (3.0 ml/column). After the addition of ScintiVerse BD (Fisher Scientific, Fair Lawn, NJ) radioactivity was quantified with a Beckman scintillation counter. Data shown are the average of duplicate determinations and were repeated at least three independent times.
Calcium Flux Assay.
Stably transfected CHO cells were plated in 96-well, black-walled culture plates (Corning Costar, Cambridge, MA) at a density of ∼5 × 104 cells/well in complete F-12 media 1 day prior to the experiment. Cells were then washed twice with calcium flux assay buffer (Hanks' balanced salt solution, 20 mM HEPES (pH 7.6), 2.5 mM probenecid, and 0.1% BSA) and loaded with 1 mM Fluo-3-AM, prepared in 10% pluronic acid (Molecular Probes, Eugene, OR) in calcium flux assay buffer for 1 h at 37°C. Cells were then washed twice with calcium flux assay buffer before analysis. Real-time calcium flux was measured in all 96 wells simultaneously for 3 min with a fluorescent imaging plate reader (FLIPR) (Molecular Devices, Sunnyvale, CA); test peptides were added 10 s into the experiment. Data were expressed as arbitrary florescence units versus time.
Immunoblot Analysis.
Lysates from COS-7 cells transiently transfected with mutant MCH receptor cDNAs were solubilized in SDS sample buffer and resolved by SDS-polyacrylamide gel electrophoresis, using 10% acrylamide Tris-glycine gels (Novex, San Diego, CA). The resolved proteins were then transferred onto a nitrocellulose membrane, using the Xcell II Blot Module apparatus (Novex). Nonspecific antibody binding was blocked by preincubating the membranes for 1 h at room temperature with 5% nonfat dry milk in 20 mM Tris (pH 7.4)/137 mM NaCl [Tris-buffered saline (TBS)] with 0.1% Tween-20. Membrane blots were then incubated overnight with primary antibody, anti-HA mouse monoclonal antibody, clone 12CA5 (Roche Molecular Biochemicals), in TBS/0.1% Tween-20/5% nonfat dry milk, followed by three washes with TBS/0.1% Tween-20. Blots were subsequently incubated with secondary antibody, anti-mouse Ig, horseradish peroxidase-linked F(ab′)2 fragment (Amersham Pharmacia Biotech, Arlington Heights, IL) in TBS/0.1% Tween-20/5% nonfat dry milk at room temperature for 1 h, followed by three washes with TBS/0.1% Tween-20. Chemiluminescence was used to detect the specifically recognized proteins with the Luminol reagent system (New England BioLabs, Beverly, MA). Light emission was observed on autoradiographic film, which was developed with a Kodak RP X-OMAT processor (Rochester, NY).
Cell Immunostaining and Flow Cytometry.
Stably transfected wild-type and mutant MCH receptor CHO cell lines were harvested with nonenzymatic cell dissociation buffer (Life Technologies) and resuspended in PBS containing 1% BSA and 0.1% sodium azide. Approximately 1 × 106 cells in 100 μl were incubated with a primary antibody, monoclonal anti-HA antibody, clone 12CA5 (Roche Molecular Biochemicals), at 10 μg/ml, on ice for 30 min. The cells were washed twice with ice-cold PBS and incubated with a secondary antibody, sheep F(ab′)2 anti-mouse IgG, coupled toR-phycoerythrin (Sigma, St. Louis, MO), at 10 μg/ml, on ice for 30 min. These cells were washed twice with ice-cold PBS and subjected to flow cytometry, using a FACS Vantage instrument (Becton Dickinson, San Jose, CA), and analyzed for cell surface staining. Specific staining was determined by parallel incubations in the absence of primary antibody.
Results
Molecular Modeling of the MCH Peptide/Receptor Complex.
The molecular model of the human MCH peptide ligand and the helical model of the human MCH receptor were constructed separately, then docked together as described in Materials and Methods. The receptor model contains two discrete hydrophilic pockets, one between transmembrane domains 3 and 7 and the other between transmembrane domains 4, 5, and 6, which are separated by a central hydrophobic region within the transmembrane bundle. This arrangement suggests potential binding site domains for the MCH peptide ligand. In this model, two alternative orientations for the MCH peptide could be proposed, depending on the hydrophilic pocket in which Arg11 and Arg14 of the ligand were placed. After attempts to manually dock MCH into the receptor in the two potential orientations, the most sterically favored appeared to be that with Arg11 facing transmembrane domains 3 and 7 and Arg14 facing transmembrane domains 4, 5, and 6, with Val12interacting with the central hydrophobic region. We then constructed the extracellular loops of the receptor about the MCH docked helical receptor model as described in Materials and Methods. Subsequent manual adjustments and molecular mechanics minimization refinements were conducted, resulting in a model of the MCH peptide-receptor complex (Fig. 1). Using this model, we identified candidate sites of interaction between the receptor and the ligand (shown in red) and tested the proposed docked orientation by site-directed mutagenesis.

Molecular model of the MCH/receptor complex. The human MCH peptide ligand represented by a blue backbone with green side chains (S, yellow; N, dark blue; O, red) is shown docked into the transmembrane helical domain pocket of the MCH receptor, represented as a black ribbon diagram. The point mutations made in this study are identified in red along the MCH receptor ribbon, which line the putative MCH binding pocket.
Site-Directed Mutagenesis of the MCH Receptor.
Melanin-concentrating hormone is a 19-amino acid peptide ligand that presumably makes multiple contacts with its receptor. The receptor-ligand model shown in Fig. 1 predicts that approximately half the ligand, including the entire cyclic ring between Cys7 and Cys16, lies within the transmembrane domains of the receptor. Based on this model, multiple candidate binding site residues were identified in the receptor protein. Eleven of these, as labeled and indicated in red on the receptor ribbon model in Fig. 1, were individually mutated to Ala and analyzed for agonist binding affinity and functional activity. We then refined the mutagenesis strategy by using other amino acids to replace the receptor residues of interest to investigate in more detail the structural requirements of the agonist binding domain.
Expression Analysis of Mutant MCH Receptors.
To determine whether the mutations made in the MCH receptor affected expression of the receptor protein, the mutations were made in receptors tagged at the amino terminus with an HA epitope. Initial immunoblot analysis of COS-7 cells transiently transfected with the HA epitope-tagged wild-type and mutant MCH receptor cDNAs confirmed the expression of the mutant receptor proteins (data not shown).
To measure cell surface expression, CHO cells were stably transfected with HA-tagged wild-type and mutant MCH receptor cDNAs. After an initial G418 (Geneticin) selection, individual clonal cells were obtained, and surface expression of the HA epitope was measured in the stable cell lines by FACS analysis, usingR-phycoerythrin-labeled HA antibody. As shown in Fig.2, nontransfected CHO cells (Fig. 2A) and the non-HA-tagged MCH receptor transfected CHO cells (Fig. 2B) displayed only background staining, whereas CHO cells transfected with the HA-tagged MCH receptor (Fig. 2C) showed a high degree of cell staining, as demonstrated by the large rightward shift in the fluorescent peak.

Immunostaining and flow cytometry of MCH receptor-expressing CHO cells. Stably transfected CHO cells were incubated with 10 μg/ml of primary antibody (anti-HA antibody) for 30 min, washed, then stained for 30 min with secondary antibody [R-phycoerythrin-coupled sheep F(ab′)2 anti-mouse IgG], washed, and then subjected to flow cytometry. Cell number is plotted against fluorescence intensity. Specific staining is represented by the black shaded curves and nonspecific staining caused by autofluorescence of the cells (incubation in the absence of 1° antibody) is represented by the light-gray shaded curves. A, nontransfected CHO cells. B, MCH receptor-transfected CHO cells. C, HA-tagged MCH receptor-transfected CHO cells. D, HA-tagged D123A mutant MCH receptor-transfected CHO cells.
Characterization of Wild-Type and Mutant MCH Receptors.
CHO cells stably transfected with wild-type MCH receptor and HA-tagged wild-type MCH receptor were compared; both bound125I-MCH with high affinity, with calculatedK d values of 3.4 and 3.1 nM, respectively (Table 1). MCH was able to activate both wild-type and tagged receptors transiently expressed in COS-7 cells, as assessed by stimulation of inositol phosphate production, with EC50 values of 18.5 ± 6.4 nM and 30.0 ± 12.8 nM, respectively (Table 2). Of the 11 HA-tagged MCH receptor mutants analyzed, most displayed modest decreases in binding affinity (a less than 2-fold increase in IC50) and in functional activation of inositol phosphate production (3- to 10-fold), compared with the wild-type MCH receptor (Table 2). However, replacement of Asp123(3.32) with Ala (D123A) resulted in a complete absence of detectable 125I-MCH binding at radioligand concentrations up to 10 nM. In addition, this mutant receptor, when stably transfected into CHO cells, was unable to stimulate inositol phosphate production, even at MCH concentrations of up to 10 μM (Table 1). Cell surface expression of the HA epitope on this mutant receptor was confirmed by antibody staining of the stable CHO cell line transfected with HA-tagged D123A receptor and by FACS analysis, which exhibited fluorescent cell staining similar to that of the HA-tagged MCH wild-type receptor (Fig. 2D). The absence of any detectable interaction of this mutant receptor with MCH, despite cell surface expression, suggested that Asp123(3.32) might play a specific role in agonist binding. To examine the role of this residue more closely, additional mutant HA-tagged MCH receptors were made and stably transfected into CHO cells, all of which were positive for HA epitope cell surface staining. Membrane saturation radioligand binding studies of D123E, D123R, D123K, and D123S HA-tagged mutant MCH receptors failed to demonstrate any125I-MCH receptor binding to these mutant receptors. Of particular interest was the observation that the relatively conservative substitution of Glu for Asp123(3.32) (D123E) was unable to support the interaction of the receptor with MCH.
Binding affinity and B max determinations from 125I-MCH saturation binding of stably transfected CHO cell membranes
125I-MCH/MCH competition binding and agonist-induced inositol phosphate production analysis of MCH receptor mutants transiently expressed in COS-7 cells
To study these mutant MCH receptors further, we employed a calcium flux assay using a FLIPR instrument. In this sensitive functional assay, both the MCH receptor and the HA-tagged MCH receptor stably expressed in CHO cells displayed robust Ca2+ responses in response to 1 μM MCH, with the EC50 value for the HA-tagged MCH receptor determined to be 6.9 ± 0.6 nM. We noted a difference in the maximal Ca2+ response between the wild-type and HA-tagged MCH receptor (Fig.3), reflecting theB max difference determined in each of these clonal cell lines (Table 1). None of the Asp123(3.32)-substituted mutant receptors elicited an agonist-induced calcium flux in response to 1 μM MCH (Fig. 3). However, transfection of cDNA encoding the wild-type MCH receptor into the clonal CHO cell lines expressing these mutant MCH receptors resulted in cells that were responsive to MCH in an inositol phosphate production assay, demonstrating that these stable cell lines are able to support MCH receptor-mediated signal transduction (data not shown). The observation that the D123A receptor is unable to mediate MCH-stimulated Ca2+ flux is consistent with the absence of MCH binding and the lack of MCH-stimulated inositol phosphate production by the mutant receptor and implies that Asp123(3.32) is critical for both ligand binding and receptor activation.

Calcium flux assay of Asp123(3.32)-substituted MCH receptor mutants. Mutant HA-tagged MCH receptor stably transfected CHO cells, as indicated, were loaded with Fluo-3-AM 1 h prior to the experiment. Real-time calcium flux was measured as fluorescence change, in arbitrary units, using a FLIPR instrument, during which MCH was added to a final concentration of 1 μM at 10 s into the 3-min experiment. Data points were collected every second for the first minute and every 6 s until the end of the experiment. Each response curve is representative of an experiment repeated at least twice in triplicate.
Characterization of MCH Peptide Analogs.
Having identified Asp123(3.32) in the MCH receptor as a critical residue for agonist interaction, we next analyzed the MCH peptide to determine potential sites of interaction with the receptor. Based on the molecular model, there are two basic Arg residues in the peptide ligand (Arg11 and Arg14; see above) that, depending on the orientation of the ligand in the binding pocket, could potentially interact with the acidic side chain of Asp123(3.32). The docked orientation shown in Fig. 1 suggests the side chain of Arg11 as the most likely candidate for this interaction. To assess the validity of the MCH-receptor complex model, and to explore the optimal orientation of the docked ligand, we synthesized analogs of the MCH peptide in which either Arg11 or Arg14was replaced with Ala and assessed them for receptor binding affinity and activation.
Competition binding assays were used to determine IC50 values for [Ala11]-MCH and [Ala14]-MCH at the wild-type receptor (non-HA-tagged). As expected, MCH was shown to be a potent inhibitor of125I-MCH binding to the receptor, with an IC50 value of 1.5 nM (Table3). When the two peptide analogs were tested for the inhibition of 125I-MCH binding, [Ala14]-MCH was equipotent to the native peptide, with an IC50 value of 2.7 nM. In sharp contrast, [Ala11]-MCH displayed a 3000-fold decrease in binding affinity for the receptor, with an IC50 value of 5.6 μM. The ability of these peptide analogs to stimulate inositol phosphate production in CHO cells stably expressing the wild-type MCH receptor was then analyzed. Both MCH and [Ala14]-MCH produced a robust inositol phosphate response, whereas [Ala11]-MCH displayed no measurable inositol phosphate production at concentrations up to 100 μM (data not shown). None of the peptides were active in stimulating inositol phosphate production in nontransfected CHO cells at concentrations up to 1 or 10 μM, demonstrating the specificity of the response.
Binding affinity determinations from 125I-MCH/MCH competition binding in stably transfected MCH receptor CHO cell membranes
To analyze the structural requirements of the side chain at position 11 of the MCH peptide in more detail, the following additional MCH analogs were synthesized: [Asp11]-MCH, [Glu11]-MCH, [Lys11]-MCH, [Asn11]-MCH, [Gln11]-MCH, and [d-Arg11]-MCH. Of these, only [Lys11]-MCH and [d-Arg11]-MCH displayed measurable inhibition of 125I-MCH binding (IC50 = 197 nM and 5.1 μM, respectively) (Table3). None of the other peptide analogs inhibited125I-MCH binding at concentrations up to 10 μM.
These peptide analogs were then tested for their ability to activate the MCH receptor (non-HA-tagged), in a calcium flux assay with a FLIPR instrument. [Ala14]-MCH gave a response equal to that of the native MCH peptide (Table 3). In contrast, [Lys11]-MCH acted as a partial agonist in this functional assay, reaching only a 67% maximal response compared with native MCH (Table 3). Additional analysis of the dose-response relationship of this interaction showed the potency of [Lys11]-MCH to be greatly reduced compared with MCH, with an EC50 value of 5.1 μM versus 12 nM for native MCH, and confirmed the partial agonist nature of the interaction (Fig. 4). None of the other peptide analogs gave any measurable Ca2+response at concentrations up to 10 μM, consistent with their inability to stimulate inositol phosphate production.

Dose-response relationship of MCH and MCH peptide analogs in a Ca2+ flux assay. MCH receptor-transfected CHO cells were loaded with Fura-2-AM 1 h prior to the experiment, and the calcium flux peak height was measured as fluorescence change, in arbitrary units using a FLIPR instrument. MCH peptide analogs [MCH (○), [Lys11]-MCH (⋄), and [d-Arg11]-MCH (△)] were added at the specified concentrations 10 s into the experimental time course of 3 min. Each experiment was performed in triplicate and repeated at least twice.
Also shown in Fig. 4 is a dose-response curve for [d-Arg11]-MCH, which caused no detectable Ca2+ flux response, even at concentrations as high as 20 times its binding IC50 value. To assess whether this peptide was an antagonist of the MCH receptor, the ability of MCH to stimulate Ca2+ flux was assessed in the presence of increasing concentrations of [d-Arg11]-MCH. The addition of 100 μM [d-Arg11]-MCH, the maximal concentration solubility allowed, resulted in a rightward shift of the MCH dose-response curve, with no decrease in the maximal response (Fig.5). The apparent EC50 for MCH activation was increased from 6 nM to 27 nM in the presence of this concentration of [d-Arg11]-MCH, demonstrating that [d-Arg11]-MCH is a competitive antagonist of the MCH receptor. Schild transformation of these data gives a calculated K b of 15.8 ± 16.2 μM for d-Arg11 MCH, consistent with the IC50 value of 5.1 μM determined for the binding of this peptide analog to the MCH receptor.

Inhibition of MCH calcium flux dose responses by [d-Arg11]-MCH. MCH receptor-transfected CHO cells were loaded with Fura-2-AM 1 h prior to the experiment, and the calcium flux peak height was measured as fluorescence change in arbitrary units using a FLIPR instrument. Ca2+ flux MCH dose responses were measured in the absence (○) or presence (●) of 100 μM [d-Arg11]-MCH. Each experiment was performed in triplicate and repeated at least twice.
Discussion
Investigation of the structural basis for the binding of MCH to its receptor suggests a critical role for Asp123(3.32) in the MCH receptor protein in both agonist binding and receptor function. In addition, we have identified Arg11 in MCH to be important for the function of the peptide as a ligand for the receptor. This residue is also important for receptor activation, as substitution by Lys results in a partial agonist, and substitution by d-Arg results in a competitive antagonist of the MCH receptor. The chemical natures of the side chains of these residues, the acidic Asp in the receptor and the basic Arg in the ligand, suggest the potential for a direct interaction between these residues, forming an ion pair that is essential for receptor activation.
An Asp residue at the position corresponding to Asp123(3.32) in the third transmembrane domain of the MCH receptor is also conserved in several other G protein-coupled receptors. This residue is absolutely conserved in all receptors whose ligands are biogenic amines, where it is thought to provide the counterion for the protonated amine group of the ligand (Strader et al., 1987). Mutagenesis studies have implicated this residue in the binding and activation of numerous G protein-coupled receptors by biogenic amines and in the antagonist-mediated blockade of receptor activation (Fraser et al., 1989; Wang et al., 1991, 1993; Gantz et al., 1992; Ho et al., 1992; Mansour et al., 1992; Ohta et al., 1994; Javitch et al., 1995; Page et al., 1995; Schwarz et al., 1995). In addition, an Asp is conserved at this position in some families of peptide receptors, including the opioid and somatostatin receptors. In the peptide receptors for which the role of this Asp has been investigated [μ- and δ-opioid (Surratt et al., 1994; Befort et al., 1999), somatostatin 2 and 3 (Nehring et al., 1995; Strnad and Hadcock, 1995), N-formyl (Prossnitz et al., 1995), and ACTH (Naville et al., 1996a,b)], it has been shown to be an important determinant of ligand binding, with effects of substitution of this residue ranging from complete receptor inactivation for most of these receptors to ligand-independent, constitutive activation for the δ-opioid receptor (Befort et al., 1999). In addition, the corresponding ACTH receptor mutation (D107N) is responsible for hereditary glucocorticoid deficiency syndrome (Naville et al., 1996b), suggesting that pathologies due to mutation of this Asp may exist in other receptor systems. The identification of Asp123(3.32) in the third transmembrane domain of the MCH receptor as a critical residue for receptor activation confirms the importance of this acidic residue frequently conserved in G protein-coupled receptor function. Interestingly, MCH is the largest ligand shown to date to interact with this conserved transmembrane Asp residue, indicating that this mechanism, initially thought to be specific to small biogenic amine ligands, is also able to mediate the activity of much larger peptide ligands.
In the present study, we have also explored the structural requirements of the MCH peptide for its activation of the MCH receptor. Previous studies on the structure-activity relationship of MCH have been limited to the fish heptade- capeptide homolog of MCH (Asp-Thr-Met-Arg-Cys-Met-Val- Gly-Arg-Val-Tyr-Arg-Pro-Cys-Trp-Glu-Val) and used in vitro bioassays of melanin concentrating and dispersing activity in fish scales. In these studies the importance of the disulfide bond for activity was demonstrated by chemical reduction (Kawazoe et al., 1987) and by the synthesis of ring contraction analogs (Lebl et al., 1988), both of which eliminated ligand activity. Additional chemical modification of either the Tyr or Arg residues significantly reduced activity (Kawazoe et al., 1987). Truncation at either the carboxy or amino terminus resulted in no loss of activity, with the minimum peptide analog retaining the potency of the native peptide being MCH(5–15) (Matsunaga et al., 1989). The identification in the present study of Arg11 as a requisite residue for binding of human MCH to its receptor is consistent with the previous data, suggesting the importance of the cyclic core of MCH as the major pharmacophore of MCH receptor function. Our data also suggest that the orientation of the basic side chain at position 11 is critical for MCH-mediated receptor activation.
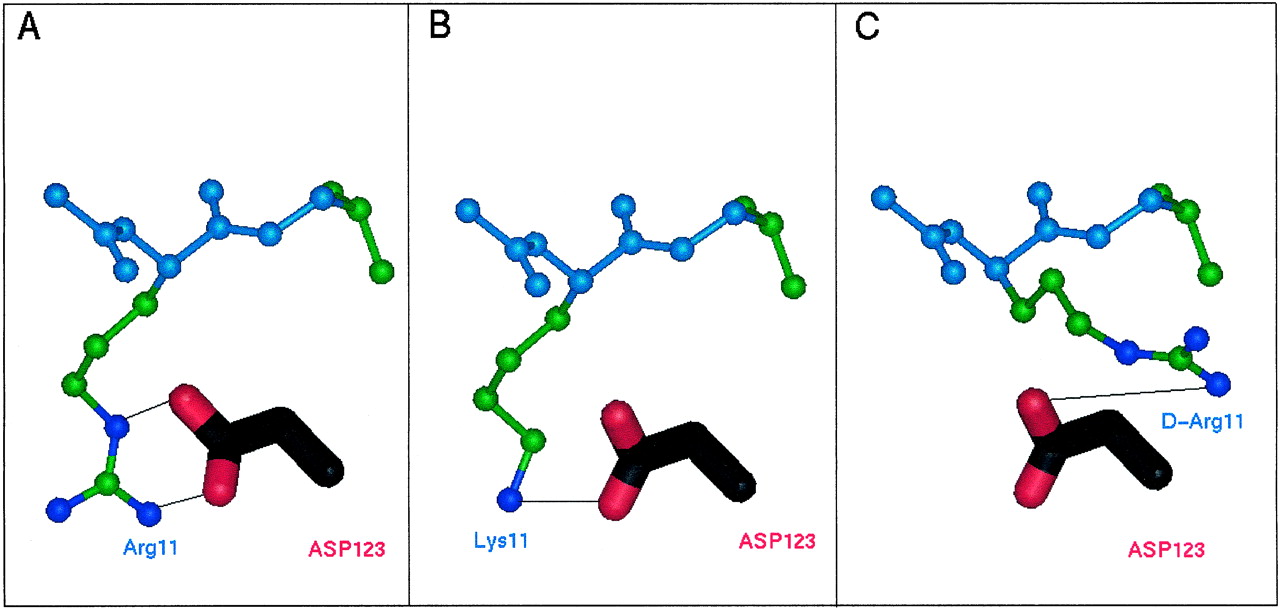
The observation that [Lys11]-MCH is a partial agonist with a 100-fold reduction in affinity compared with native MCH suggests that the proximity of the basic nitrogen in the side chain of residue 11 of the peptide to the carboxylate side chain of Asp123(3.32) on the receptor is an important parameter of both binding and activation. Further support for this notion comes from the observation that [d-Arg11]-MCH is a full competitive antagonist of the MCH receptor. Based on these observations and on the molecular model shown in Fig. 1, we propose that an interaction between Asp123(3.32) in the receptor and the side chain of Arg11 in the peptide agonist is critical for high-affinity ligand binding and receptor activation. A molecular basis for the reduced function of the peptide analogs [Lys11]-MCH and [d-Arg11]-MCH is proposed in Fig.6. In Fig. 6A, Arg11 of MCH is postulated to bind to the receptor via an ion pair between its terminal amino group and one of the carboxylate oxygens in the side chain of Asp123(3.32), plus a hydrogen bond between the Arg11 Nε and the other carboxylate oxygen. Lack of any detectable binding of [Gln11]-MCH to the receptor suggests that the ionic interaction is the more important of the two, because Gln11 would retain the potential to form a hydrogen bond at this position. In Fig. 6B, [Lys11]-MCH is able to form an ion pair with Asp123(3.32) but lacks the additional hydrogen bond of the native ligand, consistent with its partial agonist activity and 130-fold reduction in affinity. In Fig. 6C, [d-Arg11]-MCH shows the opposite stereochemistry at the binding site. In this orientation, the guanidino moiety is too far from the Asp123(3.32) side chain to allow strong binding interactions, consistent with the low-affinity, antagonist properties of [d-Arg11]-MCH.

Structural hypothesis for the function of MCH peptide analogs: molecular model of the interaction of Asp123(3.32) in the MCH receptor (black and red) with position 11 of the MCH peptide analogs (represented in ball-and-stick form; the peptide backbone is in light blue and the residue is in green; N = dark blue). Solid black lines represent postulated bond interactions (A and B) or the distance that fails to make a strong interaction (C). A, native MCH (Arg11). B, the partial agonist [Lys11]-MCH. C, the antagonist [d-Arg11]-MCH.
Overall, these data are consistent with a direct interaction between the side chains of Asp123(3.32) in the receptor and Arg11 in MCH as a key requirement for agonist-mediated receptor activation. The proximity and stereochemical nature of these interactions appears to be remarkably specific, as we were unable to restore function by switching the charged residues between the ligand and receptor; e.g., [Asp11]-MCH was not able to activate the D123R or D123K mutant receptor (data not shown). The stringent spatial requirements of the putative Arg11:Asp123(3.32) interaction for ligand binding and receptor activation would also explain the complete loss of activity of the D123E mutant MCH receptor.
Molecular modeling has suggested several other positions in the MCH receptor that may also play a role in the interaction with the MCH peptide (Fig. 1). Several of these were mutated and found to exert moderate effects on receptor function (Table 2). Of particular interest is a cluster of polar residues [Ser171(4.53), Ser174(4.56), Thr209(5.39), Gln212(5.42), and Gln276(6.55)] in transmembrane helices 4–6 of the receptor. Substitution of each of these residues with Ala results in a modest (three- to eightfold) decrease in the potency of activation by MCH. As shown in Fig. 1, the side chains of these residues would be postulated to form a network of hydrogen bonds, forming a polar cavity within the transmembrane core of the receptor. The role of this network in receptor activation remains to be determined. The molecular model shown in Fig. 1 suggests that this polar cavity might provide a potential binding site for the charged Arg14 of MCH. If this hypothesis is correct, then the observation that [Ala14]-MCH retains the agonist efficacy and binding affinity of the native MCH requires the major role of this postulated hydrogen bonding network to be to sterically permit the critical Arg11:Asp123(3.32) interaction, rather than to contribute additional binding energy through direct interactions with the ligand. Alternatively, this region might interact with a different portion of the MCH peptide. Further experiments utilizing multiple amino acid substitutions will be necessary to define the role of these residues in the interaction of the receptor with MCH.
In summary, the results of the present study are consistent with an interaction between Arg11 of MCH and Asp123(3.32) of the MCH receptor as a key component of agonist-mediated receptor activation. Further biophysical and genetic analysis will be required to completely map the interaction of MCH with its receptor.
Acknowledgments
We thank Dr. Brian Hawes and Steve Fried for collaborating in binding and functional assay development, and Xiaping Zhang for assistance with the initial immunoblot assays.
Footnotes
- Received November 23, 1999.
- Accepted March 15, 2000.
-
Dr. Douglas Macdonald, CNS/CV Research, Schering-Plough Research Institute, K-15-3-3600, 2015 Galloping Hill Rd., Kenilworth, NJ 07033. E-mail:douglas.macdonald{at}spcorp.com
Abbreviations
- Asp123(3.32)
- aspartic acid at position 123 of the MCH receptor and at the 3.32 locus in transmembrane 3 (indexing nomenclature: Ballesteros and Weinstein, 1995)
- CHO
- Chinese hamster ovary
- d-Arg
- dextrorotatory arginine
- FACS
- fluorescence analysis cell sorting
- FLIPR
- fluorescent imaging plate reader
- TBS
- Tris-buffered saline
- HA
- hemagglutinin
- WT
- wild-type
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}