


Abstract
Using a genomics-based reverse pharmacological approach for screening orphan G-protein coupled receptors, we have identified and cloned a novel high-affinity histamine receptor. This receptor, termed AXOR35, is most closely related to the H3 histamine receptor, sharing 37% protein sequence identity. A multiple responsive element/cyclic AMP-responsive element-luciferase reporter assay was used to identify histamine as a ligand for AXOR35. When transfected into human embryonic kidney 293 cells, the AXOR35 receptor showed a strong, dose-dependent calcium mobilization response to histamine and H3 receptor agonists including imetit and immepip. Radioligand binding confirmed that the AXOR35 receptor was a high-affinity histamine receptor. The pharmacology of the AXOR35 receptor was found to closely resemble that of the H3 receptor; the major difference was that (R)-α-methylhistamine was a low potency agonist of the AXOR35 receptor. Thioperamide is an antagonist at AXOR 35. Expression of AXOR35 mRNA in human tissues is highest in peripheral blood mononuclear cells and in tissues likely to contain high concentrations of blood cells, such as bone marrow and lung. In situ hybridization analysis of a wide survey of mouse tissues showed that mouse AXOR35 mRNA is selectively expressed in hippocampus. The identification and localization of this new histamine receptor will expand our understanding of the physiological and pathological roles of histamine and may provide additional opportunities for pharmacological modification of these actions.
Histamine is known to produce effects on multiple organ systems, including contraction of smooth muscle, stimulation of gastric acid secretion and cardiac contractility, and inhibition of autonomic transmitter release. Three distinct G-protein-coupled receptors, designated H1, H2, and H3, have been identified for histamine. Although the existence of these receptors has been established pharmacologically for 20 years, the H1 and H2 receptors were cloned in 1991 (Gantz et al., 1991; Yamashita et al., 1991), and the H3 receptor remained unidentified until last year (Lovenberg et al., 1999). Interestingly, the genes encoding the H1, H2, and H3 receptors share less protein sequence identity with each other than with other biogenic amine receptor family members, suggesting that these histamine receptors evolved from different ancestor sequences. Although lacking in significant overall sequence homology, the histamine receptors apparently acquired crucial elements for the recognition of histamine during their evolution. Over the last decade, there is a growing body of pharmacological evidence suggesting the possibility of further histamine receptor heterogeneity, particularly with respect to H3-mediated responses (West et al., 1990; Raible et al., 1994; Yanai et al., 1994; Leurs et al., 1996; Harper et al., 1999).
In the present communication, we report the identification of a novel histamine receptor, AXOR35, and its initial pharmacological characterization. Our data showed that although the AXOR35 receptor has many properties in common with the H3 receptor, there are also clear differences between the two receptors in terms of their pharmacology and tissue distribution.
Experimental Procedures
Materials.
The burimamide and improgan were synthesized by colleagues in the Department of Medicinal Chemistry, SmithKline Beecham Pharmaceuticals (King of Prussia, PA). Imetit, immepip,N-α-methylhistamine (N-α-MeHA), (R)-α-methyl-histamine [(R)-α-MeHA] HTMT dimaleate (6-[2-(4-imidazolyl)ethylamino]-N-(4-trifluoromethylphenyl)heptanecarboxamide) and iodophenpropit were purchased from Tocris (Ballwin, MO). Histamine, dimaprit, thioperamide, clozapine, cimetidine, and pyrilamine were from RBI (Natick, MA).
Cloning of Human AXOR35.
Full-length cloning was performed using Marathon-ready cDNA from human bone marrow (Clontech, Palo Alto, CA). PCR was performed using gene specific primers corresponding to the putative coding sequence of the gene from an unfinished genomic sequence (GenBank accession number AC007922), and AP adaptor primers (Clontech). The complete AXOR35 cDNA was isolated from human bone marrow and leukocytes by RT-PCR using the following primers (the underlined nucleotides are start and stop codons): 5′ATGCCAGATACTAATAGCACA 3′ and 5′TTAAGAAGATACTGACCGACT 3′. The product was cloned into pCR2.1 (Invitrogen, Carlsbad, CA) and fully sequenced.
The mouse AXOR35 probe used for in situ hybridization studies was obtained by screening a mouse cDNA library at low stringency using human AXOR35 cDNA as a probe. The partial cDNA is 0.5 kilobases and 68% identical to human AXOR35. For mammalian expression and functional analysis, a full-length human AXOR35 ORF was subcloned into the pCDN expression vector (Aiyar et al., 1994).
Luciferase Reporter Assay.
Transient transfection of HEK293 cells and reporter assays were performed as described previously (Fitzgerald et al., 1999). The reporter vector contained the luciferase gene under the control of a promoter consisting of three multiple response elements (MRE) and a cAMP response element (CRE).
Radioligand Binding Assay.
Membranes were prepared according to Nambi et al. (1994) from Cos-1 cells transiently transfected with pCDN/AXOR35 or pCDN using Lipofectamine plus (Life Technologies, Gaithersburg, MD). Saturation binding was performed with 2 to 60 nM [3H]histamine (Amersham Pharmacia Biotech, Piscataway, NJ) and 22 μg of membrane protein. Histamine (10 μM) was used to define nonspecific binding. Competition assays were performed using 40 nM [3H]histamine and 9 μg of membrane protein. Ninety-percent of the total binding signal was specific. Assay buffer consisted of 50 mM Tris-HCl, pH 7.5, and 10 mM MgCl2. Assays were initiated by addition of membranes in a final volume of 50 μl. Assays were incubated for 1 h at 30°C and then vacuum-filtered and rinsed on a Brandel cell harvester through Whatman GF/C filters presoaked in 0.3% polyethylenimine.
Calcium Mobilization Assays.
HEK293 cells were transiently transfected as described previously (Fitzgerald et al., 1999) with 4 μg of the chimeric G-protein Gqi5 (Conklin et al., 1993) and 11 μg of AXOR35 or pCDN. Calcium mobilization assays were performed as described previously (Fitzgerald et al., 2000) with a minor modification. Instead of sulfinpyrazone, 2.5 mM probenecid was used to inhibit organic anion transport.
Quantitative PCR (TaqMan Analysis).
Human tissues of four different persons (two men, two women, except prostate) were generously provided by Dr. R. Ravid in Netherland's Brain Bank (Amsterdam, the Netherlands), or purchased as preprepared RNA from Biochain (San Leandro, CA), CLONTECH (Palo Alto, CA). All samples were anonymous and were obtained under conditions of informed consent. The tissue samples and RNA samples were stored at −80°C. Poly(A)+RNA was isolated, reverse transcribed, and specific gene mRNA levels were measured in each sample using TaqMan analysis as described previously (Sarau et al., 1999). Primers and probe for AXOR35 were as follows: forward primer, 5′-CTGTGTCTTATAGAACTCAACATACTGGG-3′, reverse primer, 5′-CACTAAGAACCACAGCACCC-3′ and TaqMan probe 6FAM-ACGGCCACCATCAGAGTAAACAATCTTCAAG-TAMRA
For the expression of AXOR35 in human blood cells, a panel of normalized, first-strand cDNA preparations (Human Blood Fractions MTC Panel) was purchased from CLONTECH. Each cDNA sample (0.2 ng) from this panel was tested.
Tissue Preparation and in Situ Hybridization.
Male and female adult CD-1 mice (Charles River, Raleigh, NC) were dissected and tissues were collected, embedded in Tissue-Tek OCT (optimal cutting temperature) embedding media (Sakura Finetek USA Inc., Torrance, CA), and rapid-frozen in liquid nitrogen-cooled isopentane or liquid nitrogen, then stored at −80°C until used. Cryosections (5–7 μm) were prepared at −25°C (Leica CM 3050; Leica Microsystems, Inc., Deerfield, IL) and mounted on adhesive-coated slides (Instrumedics, Inc., Hackensack, NJ). The in situ hybridization method used was a modification of the procedures of Braissant et al. (1996) and Yang et al. (1999).
Results
Cloning and Sequence Analysis of the AXOR35 Receptor.
As part of a large genomics-based program to identify and clone orphan G-protein-coupled receptors as novel drug targets (Stadel et al., 1997), we identified an unfinished human genomic sequence (GenBank accession number AC007922) encoding a putative partial orphan 7 transmembrane receptor. After the use of a series of molecular cloning techniques, a full-length cDNA was isolated from human bone marrow and peripheral blood mononuclear cells (PBMC) and named AXOR35. The cDNA encodes a 390-amino-acid polypeptide with 36.6% identity to the human H3 receptor (Fig. 1A), and only 18.9% and 18.1% identity to H1 and H2 receptors, respectively. During the preparation of this article, Oda et al. (2000) reported a human G-protein-coupled receptor with a sequence (GPRv53) nearly identical to the AXOR35 receptor, differing by only three amino acids: V138→A, R206→H and R253→Q. Our sequence matches 100% with the genomic sequence (GenBank accession number AC007922).

A, amino acid alignment of AXOR35 and histamine H1, H2, and H3 receptors. The identical amino acids are boxed, and putative transmembrane domains (transmembranes 1–7) are indicated by underlines. The nucleotide and amino acid sequences have been submitted to GenBank (accession number AF325356). B, genomic structure of AXOR35 predicted from GenBank accession number AC007922. Exon sequence in upper case and intron sequence in lower case. The sizes of exons are labeled by the numbers according to the coding sequence. The sizes of introns are estimated base on the genomic sequence. *, Size can not be determined because the genomic sequences are unfinished and unordered.
Based on the mapping information from the genomic sequence (GenBank accession number AC007922), the AXOR35 receptor gene is located on chromosome 18q11.2. Its encoded protein sequence is divided into three exons encompassing amino acids 1 to 64, 65 to 119, and 120 to 390 (Fig. 1B).
Luciferase Reporter Assay of Cells Expressing the AXOR35 Receptor.
A MRE/CRE-directed luciferase reporter assay was used to identify histamine as the natural ligand for AXOR35. This assay has previously been shown to detect responses from Gi-, Gs- or Gq-coupled receptors (Fitzgerald et al., 1999). Because the AXOR35 receptor closely resembles the H3-histamine receptor, we first tested histamine and H3 agonists for their ability to induce a functional response in the cells transiently cotransfected with the reporter gene and AXOR35 or vector control. Shown in Fig. 2, in HEK293 cells transfected with the AXOR35 receptor cDNA, histamine inhibited forskolin-stimulated luciferase activity consistent with an action mediated through a Gi-coupled receptor. Histamine had no effect on pCDN-transfected cells, demonstrating that its effect was specifically mediated via the AXOR35 receptor. The H3 selective agonists immepip and N-α-methylhistamine also inhibited luciferase activity. Serotonin, dopamine, and histidine had no significant inhibitory effect on AXOR35 cDNA-transfected cells compared with pCDN-transfected cells. Additionally, norepinephrine (10 μM) and epinephrine (10 μM) (not shown) stimulated luciferase activity to 800% and 1300% over forskolin-stimulation, respectively, in both AXOR35 and pCDN-transfected cells, indicating the presence of an endogenous β2-adrenergic receptor in HEK293 cells (Fitzgerald et al., 1999).
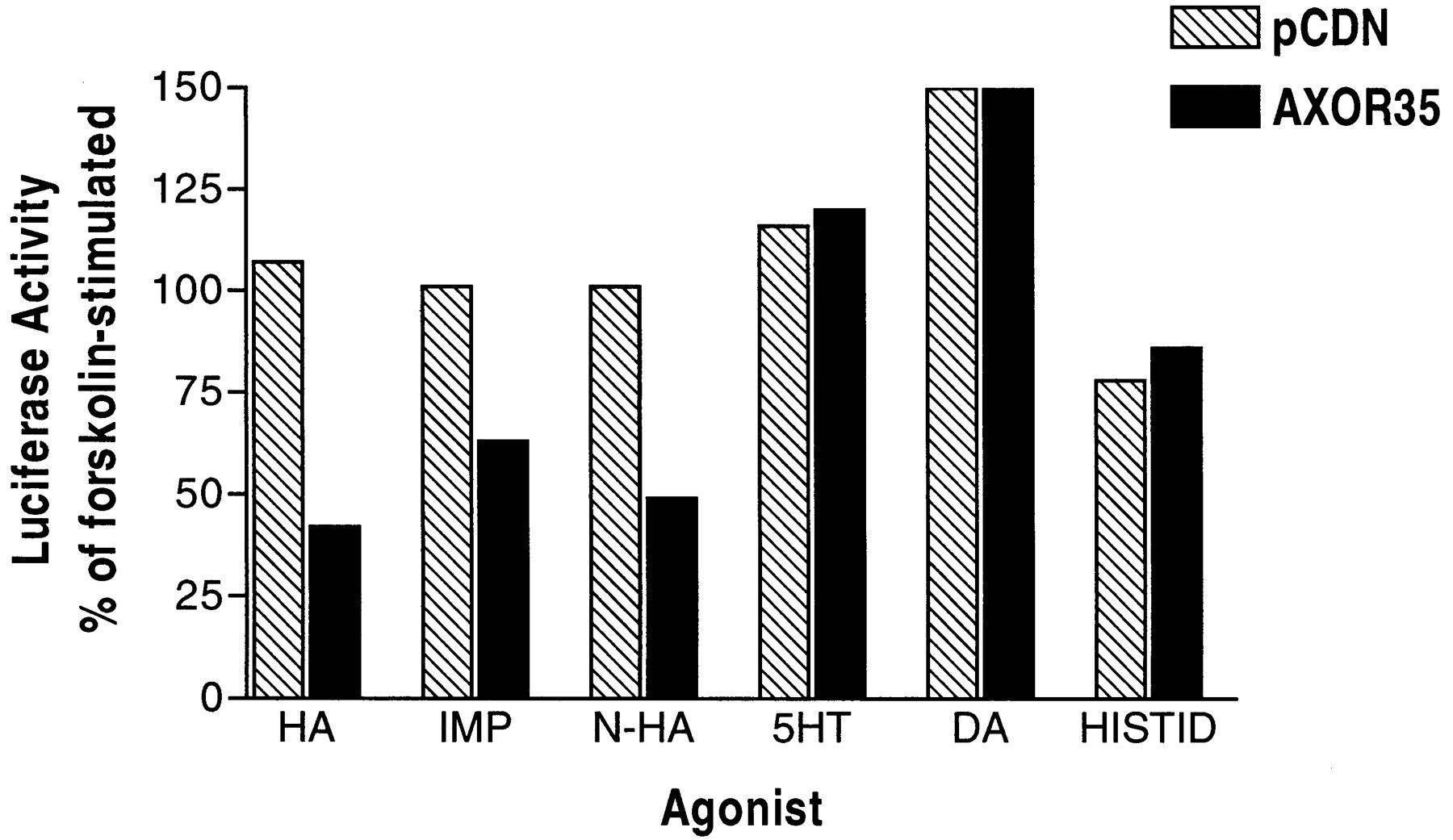
Effect of biogenic amines and H3 receptor agonists in a cAMP-driven functional assay. HEK293 cells transiently cotransfected with a MRE/CRE-luciferase reporter and AXOR35 or pCDN were treated with forskolin (5 μM) and agonists (10 μM): histamine (HA), immepip (IMP), N-α-MeHA (N-HA), serotonin (5-HT), dopamine (DA), and histidine (HISTID).
[3H]Histamine Binding Studies of the AXOR35 Receptor.
Analysis of [3H]histamine binding to membranes from COS-1 cells transiently transfected with AXOR35 cDNA showed high-affinity and saturable binding withK d and B maxvalues of 17.6 nM and 1 pmol/mg of protein, respectively (Fig.3). No binding signal was detected in membranes prepared from COS cells transiently transfected with the control vector, pCDN. Competition binding studies were performed to determine the affinities of the histaminergic agonists and antagonists. As shown in Fig. 4A, histamine and the H3-receptor agonists imetit, immepip, N-α-MeHA, and (R)-α-MeHA competed for [3H]histamine binding withK i values of 17 ± 4, 6 ± 1, 23 ± 2, 149 ± 38, and 348 ± 63 nM, respectively. The H2 agonist, dimaprit and the H1 agonist, HTMT dimaleate hadK i values of 677 ± 79 and 1229 ± 142 nM, respectively. In addition, the H3 histamine antagonists, iodophenpropit, burimamide, and thioperamide, competed for [3H]histamine binding withK i values of 18 ± 4, 160 ± 29, and 519 ± 96 nM, respectively (Fig. 4B). Clozapine, an atypical antipsychotic that blocks both 5-hydroxytryptamine2A/C and D2 dopamine receptors, showed modest affinity (K i = 693 ± 89 nM). Improgan, a novel histamine receptor agonist, which apparently does not interact with any of the known histamine receptors, had low affinity (K i = 6 μM) at AXOR 35. The H1 antagonist, pyrilamine (mepyramine) and the H2 antagonist, cimetidine, had IC50 values greater than 100 μM.

[3H]Histamine shows high-affinity and saturable binding to membranes prepared from AXOR35-transfected cells. COS-1 cells were transiently transfected with AXOR35 and cell membranes were prepared 48 h later. Saturation analysis reveals a single high-affinity and saturable binding site with aK d value of 17.6 nM and aB max value of 1 pmol/mg protein. Data are from two experiments, each of which is the mean of three samples.

Pharmacological characterization of AXOR35. A and B, competition for [3H]histamine binding. Membranes were prepared from COS-1 cells transiently transfected with AXOR35. A, the K i values (nM; mean ± S.E.M) were as follows: imetit, 6 ± 1 (●); histamine, 17 ± 4 (▪); immepip, 23 ± 2 (▴); N-α-MeHA, 149 ± 38 (○); (R)-α-MeHA, 348 ± 63 (■); dimaprit, 677 ± 79 (▿); and HTMT dimaleate, 1229 ± 142 (∗). B, the K i values (nM) were as follows: iodophenpropit, 18 ± 4 (♦); burimamide, 160 ± 29 (⋄); thioperamide, 519 ± 96 (▵); clozapine, 693 ± 89 (▾); improgan, 6000 ± 866 (+); cimetidine, >100,000 (∗); pyrilamine, >100,000 (×). Each data point is the mean of three separate experiments performed with triplicate samples. C, effect of histamine and histamine receptor agonists on calcium mobilization. HEK293 cells transiently cotransfected with AXOR35 and Gqi5were loaded with the calcium-sensitive dye fluo-3 AM. Intracellular calcium was measured using a fluorescent imaging plate reader, and the peak responses obtained after agonists addition were plotted. The EC50 values (nM; mean ± S.E.M) were as follows: imetit, 15 ± 2 (●); histamine, 19 ± 3 (▪); immepip, 29 ± 2 (▴); N-α MeHA, 56 ± 4 (○); (R)-α-MeHA, 147 ± 16 (■). Burimamide (⋄), iodophenpropit (♦), HTMT (∗), and thioperamide (▵) were tested at 10 μM. Each data point is the mean of four separate experiments performed with triplicate samples.
Calcium Mobilization Analysis of Cells Expressing the AXOR35 Receptor.
A calcium mobilization assay was established using a fluorometric imaging plate reader to obtain EC50values in response to agonist treatment. The chimeric G-protein, Gqi5 (Conklin et al., 1993), was required to permit the AXOR35 receptor to couple to calcium mobilization. The chimeric protein consisted of the Gαq subunit in which the five carboxyl-terminal residues are replaced with the corresponding carboxyl-terminal residues from Gα i2. Histamine, imetit, immepip, N-α-MeHA, and (R)-α-MeHA induced calcium mobilization responses with EC50 values of 19 ± 3 nM, 15 ± 2 nM, 29 ± 2 nM, 56 ± 4 nM, and 147 ± 16 nM, respectively, in cells cotransfected with the AXOR35 receptor and Gqi5 (Fig. 4C). HTMT (10 uM), a histamine derivative, did not significantly induce calicum mobilization in cells transfected with AXOR35 and Gqi5. The H3 antagonists, iodophenpropit, burimamide and thioperamide were tested in the calcium mobilization assay to determine whether they were behaving as agonists or antagonists at AXOR35. Iodophenpropit (10 μM) and burimamide (10 μM) showed partial agonism in AXOR35/Gqi5-transfected cells, whereas thioperamide (10 μM) showed no activity in the calcium mobilization assay. Furthermore, thioperamide dose dependently shifted the histamine dose-response curve at AXOR35 with a K Bvalue of 132 nM (r 2 = 0.94; slope = 0.78). These data demonstrate that thioperamide is acting as an antagonist at AXOR35. Cells transfected with pCDN and Gqi5 or AXOR35 receptor alone showed no response to histamine or the other compounds tested.
Expression of AXOR35 Receptor.
To investigate the physiological role of the AXOR35 receptor, we profiled the tissue distribution of this receptor in a range of human tissues using RT-PCR based TaqMan analysis (Fig. 5A). Expression of AXOR35 receptor mRNA was most abundant in PBMC, and moderately abundant in bone marrow and lung. It was expressed at a high level in human neutrophils (Fig. 5B). Interestingly, as shown in Fig.5C, the level of AXOR35 receptor expression was 3-, 11-, and 23-fold higher in resting mononuclear cells, CD4+ T cells, and CD8+ T cells than in the activated cells. A very low level of expression was detected in resting and activated CD19+ B cells and resting CD14+ monocytes.

TaqMan quantitative RT-PCR analysis of AXOR35 mRNA levels in human tissues and cell lines. A, the cDNA from the reverse transcription of 1 ng of poly(A)+ RNA from multiple tissues of four different nondiseased persons was assessed for AXOR35 mRNA and a control housekeeping mRNA, glyceraldehyde-3-phosphate dehydrogenase. Data are mRNA levels for each tissue presented as the mean ± S.E.M. of four individuals. B, the cDNA from reverse transcription of 50 ng of total RNA from primary cells or cell lines were analyzed. C. The CLONTECH human blood fractions MTC panel (0.2 ng of cDNA) was analyzed.
In situ hybridization analysis was carried out with several mouse tissues using mouse AXOR35 receptor antisense RNA as probe. Expression was detected in the hippocampal formation, particularly in the granular cell layer of the dentate gyrus and the pyramidal cell layer (Fig.6), but not in the rest of the brain and other tissues such as skeletal muscle, fat, heart, liver, spleen, thymus, lymph node, adrenal gland, spinal cord, or kidney.

In situ hybridization analysis of AXOR35 expression in mouse brain. Sense and antisense riboprobes were prepared from the 3′ half of the mouse AXOR35 coding region. A, hybridization by the antisense probe to mouse hippocampus demonstrating expression in granular layer of dentate gyrus. B, hybridization by sense probe in granular layer of dentate gyrus. C, hybridization by the antisense probe to mouse hippocampus demonstrating expression in pyramidal layer. D, hybridization by sense probe in pyramidal layer.
Discussion
Using a genomics-based approach, we identified and cloned a novel histamine receptor. Pharmacologically, the AXOR35 receptor closely resembles the H3 receptor, because selective H3 agonists like imetit and immepip are potent agonists of AXOR35. Furthermore, the mode of signal transduction seems to be identical for both H3 and AXOR35 receptors, because each couples to the Gα i G-protein subtype and subsequently inhibits adenylyl cyclase. Consistent with the Gα i coupling, histamine-induced calcium mobilization responses of the AXOR35 receptor required cotransfection with Gqi5.
The major difference noted in the pharmacology of AXOR35 and H3 histamine receptors is the low potency and affinity of (R)-α-MeHA to the AXOR35 receptor compared with the H3 receptor. This was observed in both functional (calcium mobilization) and radioligand binding assays, where (R)-α-MeHA was 8-fold less potent and 20-fold lower affinity than histamine, respectively (Figs. 3 and 5; Table 1). In contrast, (R)-α-MeHA is a potent inhibitor (IC50, 1 nM) of forskolin-induced cAMP formation in cells expressing the human H3 receptor (Lovenberg et al., 1999). Furthermore, (R)-α-MeHA had substantially higher binding affinity (K d = 0.15 nM) than histamine (K i = 3 nM) for the H3 receptor. In a variety of functional and radioligand binding assays using native H3 receptors from various animal species, (R)-α-MeHA was consistently shown to have equal or greater potency than histamine (Arrang et al., 1987; West et al., 1990) (Table 1). The low potency of this agonist in our assays clearly shows that the AXOR35 receptor has a pharmacology that differs from the H3 receptor. Very recently, Oda et al. (2000) also observed the low relative potency of (R)-α-MeHA (17- and 94- fold weaker than histamine in functional and binding assays, respectively) at a novel histamine receptor virtually identical AXOR35.
Comparison of potency/affinity of histamine (HA) and (R)-α-MeHA in a variety of functional and radioligand binding assays
Another potential difference between AXOR35 and the H3 receptor is the lower potency of thioperamide at AXOR35. Our findings in both functional (K B = 132 nM) and binding assays (K i = 519 nM) show thioperamide to have over 10-fold lower affinity than reported in functional (K B= 4 nM, Arrang et al., 1987; IC50 = 20 nM, Lovenberg et al., 1999) or radioligand binding assays (K i = 20 nM,Lovenberg et al., 1999) measuring affinity at native or recombinant H3 receptors. Assessment of the pharmacological significance of this potency difference will require the comparison of potencies for a structurally diverse series of antagonists in parallel assays using AXOR35 and H3 receptors.
Several other descriptions of H3 receptor heterogeneity (West et al., 1990; Leurs et al., 1996; Harper et al., 1999) have been reported. However, in each of these reports, the putative H3 subtypes did not differ substantially in their affinity for (R)-α-MeHA, and heterogeneity was based on differential affinity of H3 antagonists. Hence, it seems that the H3 receptor heterogeneity observed by these authors does not involve a contribution of the AXOR35 receptor. The existence of a non-H1, non-H2, and non-H3-histamine receptor has been proposed based on the ability of cimetidine and the cimetidine-like compound improgran to induce antinociception that is not mediated through known histamine receptors (Li et al., 1996, 1997). However, improgan does not stimulate AXOR35 and competes very weakly for [3H]histamine binding to AXOR35, indicating that it is not likely to be the receptor linked to antinociception. Another interesting non-H1, non-H2, or non-H3 receptor is the HTMT receptor. HTMT, a derivative of histamine, is a potent immunosuppressive agent that stimulates H1, H2, and HTMT receptors in lymphocytes (Khan et al., 1986). AXOR35, like the HTMT receptor (Qui et al., 1993), is expressed in lymphocytes and neutrophils. However, AXOR35 does not seem to be the HTMT receptor. Although HTMT has modest affinity at AXOR35, HTMT does not induce calcium mobilization in AXOR35/Gqi5-transfected cells. In addition, thioperamide, which does not antagonize the HTMT receptor (Qui et al., 1993), behaves as an antagonist at AXOR35.
Raible et al. (1994) characterized a novel histamine receptor on human eosinophils that mediated calcium mobilization. This histamine receptor had pharmacological characteristics similar to those of the AXOR35 receptor, with (R)-α-MeHA being about 30-fold less potent than histamine. Burimamide, generally regarded as an H3 antagonist, showed partial agonist activity in their assay; interestingly, we found that burimamide activates the AXOR35 receptor in our calcium mobilization assay (data not shown), providing another parallel between the native histamine receptor on human eosinophils and the AXOR35 receptor. Although burimamide was reported to have high affinity for the H3 receptor (Alves-Rodrigues et al., 1998; West et al., 1999), there is no report of agonist or partial agonist activity in H3 models. Our data showing expression of AXOR35 mRNA in peripheral blood mononuclear cells is consistent with its involvement in eosinophil calcium mobilization. Oda et al. (2000) demonstrated the expression of their novel histamine receptor in eosinophils.
The AXOR35 receptor was found to be predominantly expressed in human peripheral blood mononuclear cells, whereas the H3 receptor is expressed only in brain (Lovenberg et al., 1999). Although in situ hybridization revealed specific expression of the mouse AXOR35 receptor in restricted regions of mouse hippocampus, the in situ hybridization of the H3 receptor in rat brain showed high levels of mRNA in thalamus, ventromedial hypothalamus, and caudate nucleus (Lovenberg et al., 1999). Therefore, the distribution of the AXOR35 mRNA observed in the present study clearly differed from that of the human H3 receptor.
The expression of the AXOR35 receptor in PBMC and resting mononuclear cells, CD4+/CD8+ T cells suggests that this receptor may play a role in immune modulation. Histamine signaling through the H1 receptor is reported to have a stimulatory effect on the immune system, whereas signaling through the H2 receptor has an inhibitory effect (Beer et al., 1984; Falus et al., 1992). H3 receptor agonists were found to inhibit TNF release by stimulating the release of an anti-inflammatory cytokine, IL10 (Sirois et al., 2000). Here we identified a new histamine receptor expressed on immune cells, suggesting that some of the effects previously associated with H1, H2, and H3 histamine receptors may need to be reevaluated. The observation that AXOR35 message RNA is expressed at higher levels in resting compared with activated leukocytes is currently under further investigation.
The identification of an additional histamine receptor will help to explain many biological actions of histamine and may resolve some of the inconsistencies in reports of the pharmacology of histamine receptor agonists and antagonists. The clear pharmacological differences observed thus far between the AXOR35 and H3 receptors suggest that selective agonists and/or antagonists can be identified. With these pharmacological tools, it will be possible to differentiate the functional roles of these receptors in both normal physiological regulation and disease pathology.
Acknowledgments
We thank P. Nuthulaganti, J. J. Foley, and P. T. Buckley for many calcium mobilization studies; Dr. Paul R. Murdock and Lei Chuang for preparing TaqMan samples; and members of the sequencing core facility in the Department of Genetic Technology for DNA sequencing. We are grateful to Drs. Christine M. Debouck and David Brooks for their support and advice on this project.
Footnotes
- Received November 29, 2000.
- Accepted January 5, 2001.
-
Send reprint requests to: Dr. Yuan Zhu, Department of Molecular Biology, SmithKline Beecham Pharmaceuticals, 709 Swedeland Road, PO Box 1539, King of Prussia, PA 19406. E-mail:yuan_zhu-1{at}sbphrd.com
Abbreviations
- N-α -MeHA
- N-α-methylhistamine
- (R)-α-MeHA
- (R)-α-methylhistamine
- HTMT dimaleate
- 6-[2-(4-imidazolyl)ethylamino]-N-(4-trifluoromethylphenyl)heptanecarboxamide
- HEK
- human embryonic kidney
- MRE
- multiple responsive element
- CRE
- cAMP-responsive element
- RT
- reverse transcription/transcriptase
- PCR
- polymerase chain reaction
- PBMC
- peripheral blood mononuclear cells
- pCDN
- plasmid containing cylomegalovirus promoter, DHFB and Neo selection markers
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}