


Abstract
Cannabinoids exert most of their effects through the CB1 receptor. This G protein-coupled receptor signals inhibition of adenylyl cyclase, modulation of ion channels, and stimulation of mitogen- and stress-activated protein kinases. In this article, we report that Δ9-tetrahydrocannabinol (THC), the major active component of marijuana, induces sphingomyelin hydrolysis in primary astrocytes but not in other cells expressing the CB1 receptor, such as primary neurons, U373 MG astrocytoma cells, and Chinese hamster ovary cells transfected with the CB1 receptor cDNA. THC-evoked sphingomyelin breakdown in astrocytes was also exerted by the endogenous cannabinoid anandamide and the synthetic cannabinoid HU-210 and was prevented by the selective CB1 antagonist SR141716. By contrast, the effect of THC was not blocked by pertussis toxin, pointing to a lack of involvement of Gi/o proteins. A role for the adaptor protein FAN in CB1 receptor-coupled sphingomyelin breakdown is supported by two observations: 1) coimmunoprecipitation experiments show that the binding of FAN to the CB1 receptor is enhanced by THC and prevented by SR141716; 2) cells expressing a dominant-negative form of FAN are refractory to THC-induced sphingomyelin breakdown. This is the first report showing that a G-protein-coupled receptor induces sphingomyelin hydrolysis through FAN and that the CB1cannabinoid receptor may signal independently of Gi/oproteins.
Cannabinoids, the active components of Cannabis sativa (marijuana) and their endogenous counterparts, exert most of their central and peripheral effects by binding to specific G protein-coupled receptors (Howlett, 1995; Felder and Glass, 1998). To date, two different cannabinoid receptors have been characterized and cloned from mammalian tissues: CB1 (Matsuda et al., 1990) and CB2 (Munro et al., 1993). The CB1 receptor is mainly distributed in the central nervous system, but is also present in peripheral nerve terminals, as well as in extraneural organs such as testis, uterus, eye, vascular endothelium, spleen, and tonsils. By contrast, the expression of the CB2 receptor is almost exclusively restricted to cells and organs of the immune system. Activation of these receptors has been shown to trigger several Gi/o-protein-mediated signaling pathways. For example, both the CB1 and the CB2 receptor signal inhibition of adenylyl cyclase and stimulation of extracellular signal-regulated kinase, and the CB1 receptor is also coupled to modulation of Ca2+ and K+ channels (Howlett, 1995; Felder and Glass, 1998). The discovery of a family of endogenous ligands of cannabinoid receptors (Devane et al., 1992; Mechoulam et al., 1995) and the potential therapeutic applications of cannabinoids (Voth and Schwartz, 1997; Piomelli et al., 2000) have focused a lot of attention on cannabinoids during recent years.
One of the most intriguing and unexplored signal-transducing actions of cannabinoids is their ability to activate the sphingomyelin pathway. Thus, we have reported that Δ9-tetrahydrocannabinol (THC), the major active component of marijuana, induces sphingomyelin breakdown and intracellular ceramide accumulation in primary astrocytes (Sánchez et al., 1998b; Blázquez et al., 1999) and C6 glioma cells (Sánchez et al., 1998a; Galve-Roperh et al., 2000). Accumulating evidence shows that ceramide generated upon sphingomyelin hydrolysis acts as an ubiquitous second messenger in the regulation of many physiological events related to cellular differentiation, proliferation, and programmed death (Kolesnick and Krönke, 1998;Hannun and Luberto, 2000). The link between receptor activation, sphingomyelin breakdown, and ceramide generation is mostly supported by comprehensive studies on the 55-kDa tumor necrosis factor (TNF) receptor, the 75-kDa neurotrophin receptor, and CD95/Fas. In addition, exposure of cells to physical (e.g., ultraviolet radiation, heat shock), oxidative (e.g., reactive oxygen species), bacterial (e.g., lipopolysaccharide), or viral (e.g., human immunodeficiency virus 1) stimuli may evoke changes in the activity of the sphingomyelin cycle (Kolesnick and Krönke, 1998; Levade and Jaffrézou, 1999;Hannun and Luberto, 2000). The potential pathophysiological implications of cannabinoid-induced sphingomyelin hydrolysis, however, are hampered by the absolute lack of knowledge of the molecular mechanism responsible for this effect. The present work was therefore undertaken to study the mechanism by which cannabinoids induce sphingomyelin breakdown.
Materials and Methods
Reagents.
The following materials were kindly donated: the plasmids carrying the full-length and truncated forms of FAN cDNA (Dr. S. Adam-Klages, Kiel University, Germany); the Chinese hamster ovary (CHO) cells stably transfected with the rat CB1cannabinoid receptor cDNA (Dr. T. I. Bonner, National Institutes of Health, Bethesda, MD, and Dr. Z. Vogel, The Weizmann Institute, Rehovot, Israel); the rabbit antibody raised against residues 1 to 14 of rat CB1 cannabinoid receptor (Dr. A. Howlett, St. Louis University, St. Louis, MO); SR141716 and SR144528 (Sanofi Recherche, Montpellier, France); JWH-133 (Dr. J. W. Huffman, Clemson University, SC); and HU-210 (Prof. R. Mechoulam, Hebrew University, Jerusalem, Israel). The rabbit antibody raised against the 200 C-terminal residues of FAN was from Zymed Laboratories (South San Francisco, CA). THC, anandamide, and methanandamide were from Sigma (St. Louis, MO).
Cell Culture.
Rat cortical astrocytes (Sánchez et al., 1998b), rat cortical neurons (Sánchez et al., 1998a), the human astrocytoma U373 MG (Sánchez et al., 1998a), and CHO cells transfected with the rat CB1 receptor cDNA (Gómez del Pulgar et al., 2000) were cultured as described previously. The human umbilical-vein endothelial cell line ECV304 was cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum. Twenty-four hours before the experiment, cells were transferred to their respective serum-free media. Stock solutions of cannabinoids were prepared in dimethyl sulfoxide. Control incubations had the corresponding dimethyl sulfoxide content. No significant influence of dimethyl sulfoxide was observed on any of the parameters determined at the final concentration used (0.1%, v/v).
Cell Transfection.
ECV cells were transfected by electroporation (240 V, 960 μF) with 10 μg of pcDNA3 carrying the full-length (encoding amino acids 3–917) or a truncated, dominant-negative form (encoding amino acids 703–917) of the FAN cDNA (Adam-Klages et al., 1996; Ségui et al., 1999). Transfected cells were maintained in culture in the presence of 0.5 mg/ml G418.
Sphingomyelin and Ceramide Levels.
Cells were incubated for 48 h in their respective chemically-defined media supplemented with 1 μCi of [3H-methyl]choline (sphingomyelin levels) or 1 μCi ofl-[U-14C]serine (ceramide levels) per well. Reactions were started by the addition of the different modulators and stopped with 0.85 ml of methanol at the times indicated in the figure legends. Lipids were extracted and saponified, and ceramide and sphingomyelin were quantified (Blázquez et al., 1999).
Western Blot Analysis of FAN and the CB1Receptor.
Cells were scraped from the plates, sonicated (2 × 5 s) on ice, and the particulate fraction was obtained after centrifugation at 40,000g for 60 min. Samples were subjected to SDS-polyacrylamide gel electrophoresis and proteins were transferred onto nitrocellulose membranes. After blocking the blots with 0.5% nonfat dry milk in phosphate-buffered saline supplemented with 0.1% Tween 20, Western blots were performed with the anti-FAN (1:5000) or anti-CB1 receptor antibody (1:5000), and then with an anti-rabbit peroxidase-conjugated secondary antibody (1:5000). Blots were finally subjected to luminography with an enhanced chemiluminescence detection kit.
Coimmunoprecipitation of FAN and the CB1Receptor.
Cells were exposed to different agents. Reactions were terminated by washing cells with ice-cold PBS (10 mM NaPi, 150 mM NaCl, pH 7.4) supplemented with 20 mM NaF, 20 mM NaPPi, 1 mM NaVO4, and 5 mM EDTA, and subsequent addition of ice-cold lysis buffer consisting of 50 mM HEPES, pH 7.4, 150 mM NaCl, 10% (v/v) glycerol, 1% (v/v) Triton X-100, 50 mM NaF, 1 mM NaVO4, 10 mM β-glycerophosphate, 1 mM phenylmethylsulfonyl fluoride (PMSF), 10 μg/ml leupeptin, and 10 μg/ml aprotinin. The CB1 receptor was immunoprecipitated from cell lysates (0.5–1.0 mg of total protein) with the anti-CB1 receptor antibody (10 μg) bound to protein G-agarose (Sánchez et al., 1998b). Samples were washed five times with 50 mM HEPES, pH 7.4, and 150 mM NaCl, supplemented with the aforementioned proteinase inhibitors. Immune complexes were subjected to SDS-polyacrylamide gel electrophoresis, and Western blot analysis of FAN was performed as described above. Equal protein loading was checked by Coomasie blue staining.
Statistical Analysis.
Results shown represent the means ± SD of the number of experiments indicated in every case. Statistical analysis was performed by analysis of variance. A post hoc analysis was made by the Student-Neuman-Keuls test.
Results and Discussion
The CB1 Cannabinoid Receptor of Astrocytes Is Coupled to Sphingomyelin Hydrolysis through a Pertussis Toxin-Insensitive Pathway.
We have recently reported that THC induces sphingomyelin breakdown and intracellular ceramide accumulation in primary astrocytes in a time- and dose-dependent manner (Sánchez et al., 1998b;Blázquez et al., 1999). Because rat primary astrocytes express the CB1 receptor mRNA (Bouaboula et al., 1995a) and protein (Sánchez et al., 1998a), experiments were conducted to test whether THC-evoked sphingomyelin breakdown was dependent on this receptor. As shown in Fig. 1, the effect of 1 μM THC was prevented by 1 μM SR141716, a selective CB1 receptor antagonist, but not by 1 μM SR144528, a selective CB2 receptor antagonist. The synthetic cannabinoid agonist HU-210 at 50 nM also stimulated sphingomyelin hydrolysis. The endogenous cannabinoid ligand anandamide induced sphingomyelin breakdown at 50 but not at 25 μM. Because primary astrocytes have a high capacity to take up and degrade anandamide (Di Marzo et al., 1994), we tested the effect of PMSF, an inhibitor of anandamide hydrolysis, on anandamide action. When coincubated with 100 μM PMSF, 25 μM anandamide was able to evoke maximal sphingomyelin breakdown (Fig. 1). Likewise, methanandamide, a stable synthetic analog of anandamide, induced maximal sphingomyelin hydrolysis at 25 μM, indicating that anandamide action is blunted by cellular degradation. The synthetic CB2 selective agonist JWH-133 at 50 nM did not significantly affect sphingomyelin levels (Fig. 1). Interestingly, although it is well established that cannabinoid receptors are coupled to Gi/oproteins, 50 ng/ml pertussis toxin was unable to prevent THC-induced sphingomyelin breakdown (Fig. 1), whereas under identical experimental conditions, pertussis toxin fully blocked THC-induced activation of extracellular signal-regulated kinase (Bouaboula et al., 1995b) and protein kinase B (Gómez del Pulgar et al., 2000) (results not shown).

The CB1 cannabinoid receptor of astrocytes is coupled to sphingomyelin hydrolysis. Astrocytes were incubated with or without 1 μM SR141716 or SR144528 (30 min), 100 μM PMSF (30 min), or 50 ng/ml pertussis toxin (PTX, 14 h), and then with or without 1 μM THC, 50 nM HU-210, 25–50 μM anandamide (AEA), 25 μM methananandamide (M-AEA), or 50 nM JWH-133 for 10 additional min. Results are expressed as percentage of incubations with no additions and correspond to six different experiments. Significantly different from incubations with no additions: *P < 0.05; **P < 0.01.
To test whether THC-induced sphingomyelin hydrolysis also occurs in other cells expressing the CB1 receptor, primary neurons, U373 MG astrocytoma cells, and CHO cells transfected with the CB1 receptor cDNA were examined. However, THC was unable to induce a significant breakdown of sphingomyelin in these cells (Fig. 2).
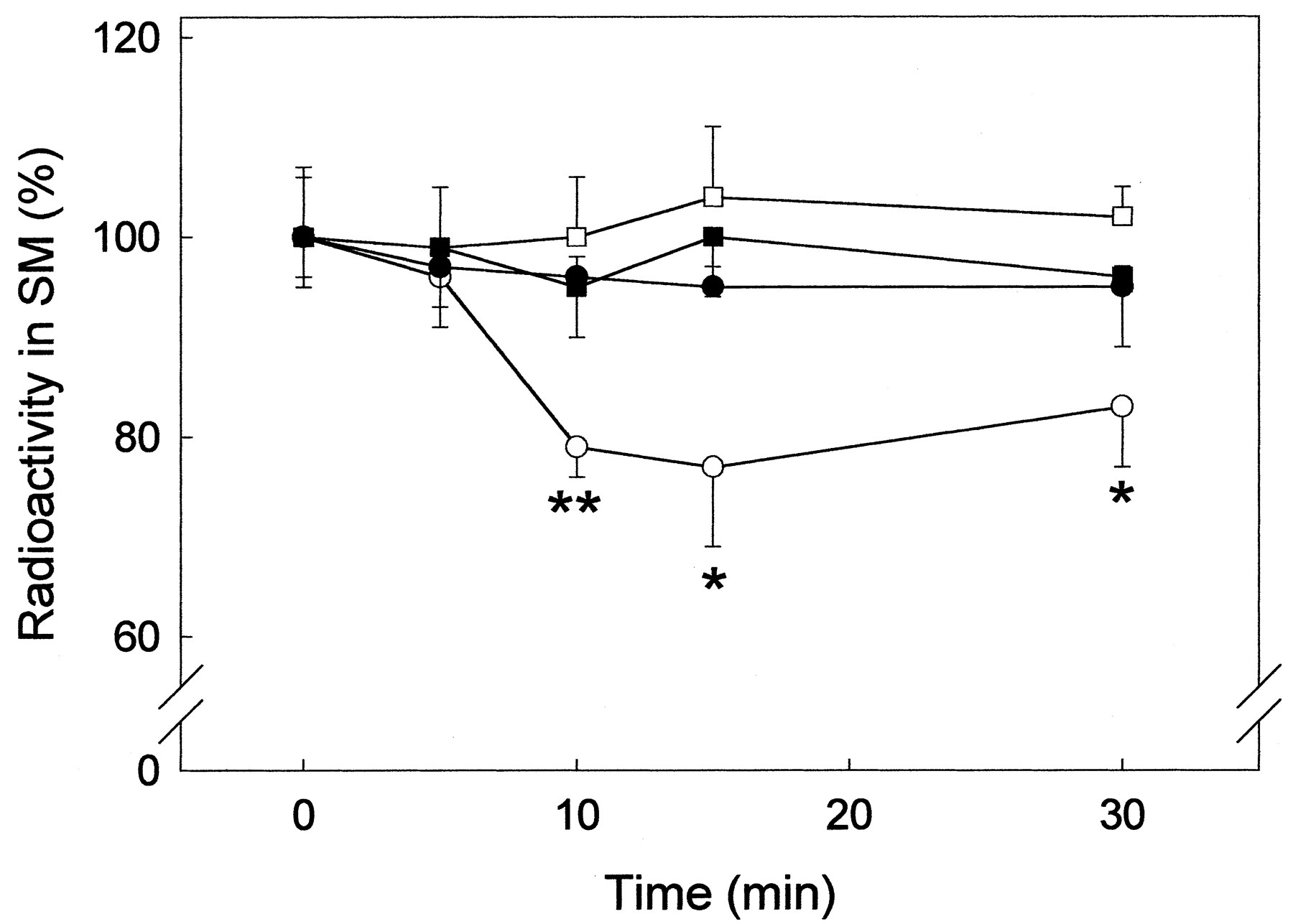
Stimulation of sphingomyelin hydrolysis by THC in astrocytes but not in other cell types. Primary astrocytes (○), U373 MG astrocytoma cells (●), primary neurons (■), and CHO cells stably transfected with the CB1 receptor (▪) were exposed to 1 μM THC for the times indicated and the radioactivity in cellular sphingomyelin (SM) was determined. Results are expressed as percentage of incubations with no additions and correspond to four different experiments. Significantly different from the corresponding incubations with no additions: *P < 0.05; **P < 0.01.
The Adaptor Protein FAN Couples the CB1 Cannabinoid Receptor of Astrocytes to Sphingomyelin Breakdown.
Extensive studies on the 55-kDa TNF receptor have finely defined the domains of the receptor that allow coupling to sphingomyelinase activation (Adam-Klages et al., 1998; Kolesnick and Krönke, 1998). A protein designated as FAN (for factor associated with neutral sphingomyelinase activation) has been shown by Krönke and coworkers (Adam-Klages et al., 1996; Kreder et al., 1999) to couple the NSD (for neutral sphingomyelinase activation domain) of the 55-kDa TNF receptor to neutral sphingomyelinase activation. FAN contains five WD repeats in its C-terminal portion that serve as functional motifs to facilitate defined protein-protein interactions. Interestingly, β subunits of heterotrimeric G proteins are also WD-repeat proteins (Adam-Klages et al., 1998). We therefore examined whether the CB1receptor may be coupled to FAN in primary astrocytes and in U373 MG astrocytoma cells, the latter selected as an example of cells in which THC is unable to induce sphingomyelin hydrolysis. For this purpose, the CB1 receptor was precipitated with a specific antibody and FAN was subsequently detected by immunoblotting. As shown in Fig. 3, FAN from primary astrocytes was precipitated by the anti-CB1 receptor antibody. Furthermore, pretreatment of astrocytes with THC favored the binding of FAN to the CB1 receptor, whereas SR141716 prevented this association. By contrast, the effect of THC was not evident in U373 MG astrocytoma cells, despite the presence of FAN in these cells.

Association of FAN to the CB1 cannabinoid receptor in astrocytes but not in U373 MG astrocytoma cells. On the left, the presence of FAN in astrocyte and U373 MG astrocytoma cell lysates is evidenced by Western blotting (WB) with the anti-FAN antibody. For coimmunoprecipitation studies, astrocytes and U373 MG astrocytoma cells were treated with THC and/or SR141716 (SR) as described in Fig. 1. Cellular lysates were immunoprecipitated (IP) with the anti-CB1 antibody and Western blots were subsequently performed with the anti-FAN antibody as described underMaterials and Methods. Representative luminograms are shown. Similar results were obtained in two other experiments.
To test whether the FAN-CB1 receptor interaction is functionally relevant, we used ECV304 cells stably transfected with either full-length FAN or a truncated, dominant-negative form of FAN that does not bind to the NSD domain of the 55-kDa TNF receptor and therefore blocks TNF-induced sphingomyelin breakdown and ceramide generation (Adam-Klages et al., 1996; Ségui et al., 1999). In line with a previous report (Liu et al., 2000), ECV304 cells were shown to express the CB1 receptor (Fig.4A). As shown in Fig. 4B, THC induced a transient breakdown of sphingomyelin in cells expressing wild-type FAN but not in cells expressing dominant-negative FAN. The involvement of the CB1 receptor in THC-induced sphingomyelin hydrolysis was proved by the antagonistic effect of SR141716. As expected, THC-induced sphingomyelin breakdown occurred in concert with an elevation of intracellular ceramide levels (Fig. 4C). HU-210 also induced sphingomyelin hydrolysis in ECV304 cells expressing wild-type FAN, and this effect was prevented by SR141716 (results not shown).

Cells expressing dominant-negative FAN are refractory to THC-evoked sphingomyelin breakdown. A, presence of the CB1 receptor in ECV304 cells expressing wild-type FAN or a dominant-negative form of FAN (ΔFAN). U373 MG astrocytoma cells were used as a control. A representative luminogram is shown. Similar results were obtained in two other experiments. B and C, ECV304 cells expressing dominant-negative (■) or wild-type FAN (▨) were exposed to 1 μM THC for the times indicated and the radioactivity in cellular sphingomyelin (SM) (B) and ceramide (C) was determined. Cells expressing wild-type FAN were also incubated with 1 μM SR141716 for 30 min followed by an additional 10 min with 1 μM THC (▧). Results are expressed as percentage of incubations with no additions and correspond to six different experiments. Significantly different from incubations with no additions: *P < 0.05; **P < 0.01.
The notion that FAN is coupled to the CB1receptor is strengthened by the homology between the NSD of the 55-kDa TNF receptor and a region of the CB1 cannabinoid receptor. NSD is a short, nine-amino-acid motif that includes the sequence DSAHK (Adam-Klages et al., 1996), and the CB1 receptor contains the highly homologous sequence DCLHK at amino acid positions 431 to 435 (Matsuda et al., 1990). The DCLHK sequence 1) is conserved in rat, human, mouse and cat CB1 receptors (see SwissProt data bank: CB1R); 2) is located in the C-terminal portion of the CB1 receptor (i.e., in a domain of the protein potentially involved in the interaction with cytoplasmic effectors); and 3) is not present in the CB2 receptor, which, in HL-60 cells at least, is not coupled to sphingomyelin hydrolysis (results not shown). It is difficult to evaluate whether the aforementioned sequence homology between the NSD of the 55-kDa TNF receptor and the NSD-like portion of the CB1receptor is actually significant. In this respect, it is worth noting that CD40, a member of the TNF receptor superfamily, has been recently shown to evoke sphingomyelin hydrolysis via FAN (Ségui et al., 1999). The sequence QETLH in the cytoplasmic domain of CD40, the one that fits better with a portion of the NSD of the 55-kDa TNF receptor (EDSAH), shares a lower similarity with the latter than the CB1 receptor. Nevertheless, as we are aware, because the CB1 receptor is not coupled to FAN in U373 MG astrocytoma cells, which express FAN, additional, still-unknown factors may be necessary for the CB1 receptor to activate sphingomyelin hydrolysis.
Conclusions
The sphingomyelin cycle plays and important role in the regulation of cell physiology in the central nervous system (Kolesnick and Krönke, 1998). Ceramide generated by challenge of astroglial cells to cannabinoids may serve as a second messenger in the control of several functions [e.g., metabolic regulation (Sánchez et al., 1998b; Blázquez et al., 1999) and induction of apoptosis (Galve-Roperh et al., 2000)]. The notion that the CB1 cannabinoid receptor, like the 55-kDa TNF receptor, may control the activity of the sphingomyelin cycle (the present report) and of mitogen- (Bouaboula et al., 1995b) and stress-activated protein kinases (Rueda et al., 2000) points to a general role of cannabinoids as modulators of glial cell fate. By showing that the CB1 cannabinoid receptor may be coupled to FAN independently of Gi/o proteins, this report opens a new conceptual view on the mechanism of cannabinoid action and contributes to the novel idea that G protein-coupled receptors may signal via ceramide (Limatola et al., 1999) as well as by mechanisms alternative to the classical heterotrimeric-G protein paradigm (Hall et al., 1999).
Acknowledgments
We are indebted to V. Garcia, N. Auge and T. Gómez del Pulgar for expert technical assistance.
Footnotes
- Received October 23, 2000.
- Accepted January 19, 2001.
-
Send reprint requests to: Dr. Manuel Guzmán, Department of Biochemistry and Molecular Biology I, School of Biology, Complutense University, 28040 Madrid, Spain. E-mail: mgp{at}bbm1.ucm.es
-
This study was supported by grants from Comisión Interministerial de Ciencia y Tecnologı́a (PM 98/0079), Comunidad Autónoma de Madrid (CAM 08.5/0017/98), and Institut National de la Santé et de la Recherche Médicale.
Abbreviations
- THC
- Δ9-tetrahydrocannabinol
- TNF
- tumor necrosis factor
- CHO
- Chinese hamster ovary
- PMSF
- phenylmethylsulfonyl fluoride
- FAN
- factor associated with neutral sphingomyelinase activation
- NSD
- neutral sphingomyelinase activation domain
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}