


Abstract
Salmeterol is a long-acting β2-adrenergic receptor (β2AR) agonist that is widely used as a bronchodilator for the treatment of persistent asthma and chronic obstructive pulmonary disease in conjunction with steroids. Previous studies demonstrated that salmeterol showed weak efficacy for activation of adenylyl cyclase; however, its efficacy in the complex desensitization of the β2AR remains poorly understood. In this work, we provide insights into the roles played by the G protein–coupled receptor kinase/arrestin and protein kinase A in salmeterol-mediated desensitization through bioluminescence resonance energy transfer (BRET) studies of liganded-β2AR binding to arrestin and through kinetic studies of cAMP turnover. First, BRET demonstrated a much reduced efficacy for salmeterol recruitment of arrestin to β2AR relative to isoproterenol. The ratio of BRETISO/BRETSALM after 5-minute stimulation was 20 and decreased to 5 after 35 minutes, reflecting a progressive decline in BRETISO and a stable BRETSALM. Second, to assess salmeterol efficacy for functional desensitization, we examined the kinetics of salmeterol-induced cAMP accumulation (0–30 minutes) in human airway smooth muscle cells in the presence and absence of phosphodiesterase inhibition. Analysis of shaping of cAMP turnover for both agonists demonstrated significant salmeterol desensitization, although it was reduced relative to isoproterenol. Using an isoproterenol rescue protocol after either short-term (10 minutes) or long-term (2 and 14 hours) salmeterol pretreatments, we found that salmeterol progressively depressed isoproterenol stimulation but did not prevent subsequent rescue by isoproterenol and additional isoproterenol-mediated desensitization. Our findings reveal a complex efficacy for functional desensitization, demonstrating that although salmeterol shows weak efficacy for adenylyl cyclase activation and G protein–coupled receptor kinase/arrestin-mediated desensitization, it acts as a strong agonist in highly amplified protein kinase A–mediated events.
Introduction
Long-acting β-adrenoceptor agonists (LABAs) such as salmeterol, properly used only in combination with inhaled corticosteroids, have shown proven efficacy as a maintenance therapy to avoid recurrence of asthmatic episodes (Cazzola et al., 2013), although controversy remains primarily based on increased morbidity and mortality for LABA use alone (Khianey and Oppenheimer, 2011). In spite of its clinical efficacy and extended action (10–12 hours), the characterization of the mechanisms of salmeterol activation and desensitization of the β2 adrenergic receptor (β2AR) has proven difficult, primarily because of its weak efficacy for β2AR stimulation of adenylyl cyclase (AC), high affinity for the β2AR (Kd = 1 to 2 nM), and extreme hydrophobicity (Rhodes et al., 1992; Clark et al., 1996; January et al., 1997, 1998). Its rapid partitioning into plasma membranes renders it highly resistant to washout, confounding facile determination of its efficacy for desensitization. Thus, many problems remain unresolved, in particular the multifaceted nature of efficacy for the complex desensitization process (Clark et al., 1999; Kenakin and Christopoulos, 2013; van der Westhuizen et al., 2014).
Agonist-mediated desensitization of the β2AR involves a highly amplified rapid G protein–dependent cAMP-dependent protein kinase A (PKA)–mediated phosphorylation of the receptor and activation of cyclic-nucleotide phosphodiesterases (PDEs), coupled with the G protein–independent pathway (January et al., 1997; Tran et al., 2004; Xin et al., 2008; Vayttaden et al., 2010). The latter is initiated by agonist-dependent G protein–coupled receptor kinase (GRK)–mediated phosphorylation of the β2AR, which in turn leads to arrestin binding to GRK-phosphorylated active β2AR and internalization (Clark et al., 1999; Tran et al., 2004). It should be noted that the GRK pathway is not strictly G protein–independent since to the extent that GRK2 and GRK3 are involved, their membrane localization requires βγ subunit binding (Lodowski et al., 2003). An additional component becomes important in extended treatment time: downregulation (t1/2 = 3 to 4 hours), which is still incompletely understood (Williams et al., 2000; Liang et al., 2008). The consequence of agonist efficacy on these processes is an additional confounding factor, since each interaction of the agonist-β2AR in protein–protein interactions (Gs, GRKs, and arrestin) must exhibit differential efficacy. When assessing agonist efficacy for β2AR desensitization, it is important to distinguish direct molecular measures of β2AR desensitization (e.g., efficacy of AC activation and GRK phosphorylation) from what we refer to as functional desensitization, the measure of downstream actions (e.g., cAMP turnover by PDE, and bronchodilation). Lack of distinction of these processes understandably has led to some confusion in defining salmeterol efficacy.
Modeling and simulations based on quantitative assessment of the rates of key parameters have provided important insights into the complexity of β2AR desensitization, with both endogenous and β2AR overexpression systems (Violin et al., 2008; Xin et al., 2008; Vayttaden et al., 2010). In our modeling of isoproterenol stimulation and desensitization of the GRK pathway, we found excellent agreement with six parameters measured after strong (isoproterenol) or weak (salmeterol) agonist stimulation: the rates of GRK site phosphorylation, dephosphorylation, internalization, recycling, desensitization, and resensitization (Vayttaden et al., 2010). For salmeterol, we found optimum validation of experimental data when we assumed that the affinity of salmeterol-β2AR for arrestin was much reduced relative to isoproterenol. This assumption was based on our prior evidence that salmeterol only weakly drives internalization and arrestin translocation (Moore et al., 2007). Simulations demonstrated that salmeterol caused decreased GRK pathway–mediated desensitization over the 0- to 30-minute time period of stimulation compared with isoproterenol, consistent with our prior experimental measurements (Clark et al., 1996; January et al., 1998; Tran et al., 2004).
In contrast with quantitative analysis of β2AR-level desensitization, defining functional desensitization based on cAMP turnover by PDE or bronchodilation is more problematic. The role of PDE in isoproterenol stimulation of cAMP accumulation in HEK293 cells expressing only endogenous receptors has also been modeled and simulated (Violin et al., 2008; Xin et al., 2008). These findings highlighted the important role of PKA activation of PDE, which was nearly equivalent to the GRK pathway in terms of its effect on the rapid phase of functional desensitization (to be distinguished from β2AR desensitization). Because of the amplification and sensitivity of the cAMP/PKA-mediated events, weak partial agonists such as salmeterol will fully activate these components of desensitization (Tran et al., 2004) even at concentrations well below their EC50 values; they thus act as strong agonists, while having minimal effects on the occupancy-dependent GRK pathway. It was shown that β2AR levels are another major factor to consider in measuring agonist efficacy (Whaley et al., 1994; January et al., 1998; Clark et al., 1999; Clark and Knoll, 2002; Xin et al., 2008; Charlton, 2009). With high β2AR expression, salmeterol appears as a strong agonist (Emax = 90–100% of isoproterenol), whereas with low endogenous β2AR levels, the Emax for salmeterol stimulation of AC is much reduced (efficacy 10–20% of isoproterenol). Furthermore, salmeterol’s rapid uptake and sequestration into plasma membranes has presented a major obstacle to quantitative assessment of desensitization by the classic washout and restimulation protocols. Whether in measurements of either cell-free membrane preparations of AC (Clark et al., 1996; January et al., 1998) or functional desensitization in intact cells (Düringer et al., 2009; Nino et al., 2009; Cooper et al., 2011), membrane-sequestered salmeterol has a profoundly confounding effect. Our studies demonstrated that short-term treatment of the β2AR with salmeterol caused much reduced desensitization compared with strong agonists in membrane assays (Clark et al., 1996; January et al., 1998). On the basis of studies comparing the effects of equieffective concentrations of salmeterol and strong agonists on functional desensitization in human airway smooth muscle (HASM) cells by measure of either cAMP accumulation or bronchodilation and bronchoprotection, it was concluded that salmeterol was a strong agonist (Düringer et al., 2009; Cooper et al., 2011).
The goal of this study was to clarify these issues concerning the efficacy of salmeterol activation and desensitization mediated by PKA regulation of PDE and the GRK pathway. To this end, we report first on studies testing our proposal that salmeterol binding to the β2AR results in reduced efficacy for its complex with arrestin. Second, we further investigated salmeterol activation and functional desensitization in HASM cells. This was accomplished through analysis of both the kinetics of isoproterenol and salmeterol stimulation of cAMP turnover with or without PDE inhibition, as well as the effects of short- and long-term salmeterol pretreatment on subsequent “rescue” by isoproterenol stimulation to facilitate parsing of the contribution of receptor-level desensitization and PDE to the overall desensitization process.
Materials and Methods
[2,8-3H]adenine was purchased from PerkinElmer (Waltham, MA). Cell culture medium was purchased from Gibco/Life Technologies (Grand Island, NY) or Corning (Manassas, VA). Salmeterol, ICI-118.551 [(±)-1-[2,3-(dihydro-7-methyl-1H-inden-4-yl)oxy]-3-[(1-methylethyl)amino]-2-butanol hydrochloride], carazolol, and isoproterenol were purchased from Sigma-Aldrich (St. Louis, MO). Salmeterol and carazolol were dissolved in dimethylsulfoxide (DMSO) and stored at 10 mM. Isoproterenol was stored at 10 mM with AT (100 mM ascorbate and 10 mM thiourea, pH 7.4; AT was diluted 1:100 for assay). ICI-118.551 was dissolved in H2O as a 10-mM solution. RR-formoterol was obtained from Sunovion Pharmaceuticals Inc. (Marlborough, MA) and dissolved in DMSO at 10 mM. Restriction endonucleases and other DNA-modifying enzymes were obtained from New England Biolabs (Ipswich, MA). The Renilla luciferase substrate coelenterazine-h was acquired from Prolume-NanoLight Technology (Pinetop, AZ). Coelenterazine-h was dissolved in 100% ethanol at a concentration of 5 mM [1000 times the final concentration used in bioluminescence resonance energy transfer (BRET) assays].
Plasmid Constructs.
The plasmid g3NVE-1 (provided by Vsevolod V. Gurevich, Vanderbilt University, Nashville, TN) containing the sequence of bovine β-arrestin 2 fused to Venus (enhanced variant of yellow-shifted fluorescent protein; Nagai et al., 2002) at its N-terminus was constructed using a modified version of the pGEM-2 in vitro transcription vector (Promega, Madison, WI), as previously described (Gimenez et al., 2012). In brief, Venus was amplified by PCR using the 5′…agtcagaattcgcgatcgcggccacgatggtgagcaagggcga…3′ forward primer, which adds EcoRI and AsiSI restriction enzyme recognition sites upstream of the start codon, and the 5′…tctcccccatggagtcgagcgctcgccgagacttaagtccggaggtggcct…3′ reverse primer, which codes for a short spacer with the “SGLKSRRALDS” sequence and an in-frame NcoI site. Venus was subcloned into the EcoRI to NcoI sites of pGEM-2. The β-arrestin 2 sequence was then subcloned in frame with the Venus-spacer sequence using NcoI and HindIII. The Venus–β-arrestin 2 fused sequence was finally ligated into a modified version of the pcDNA3 mammalian expression vector (Life Technologies, Carlsbad, CA) using EcoRI and HindIII to generate P3VEA3-1.
Renilla luciferase variant 8 (RLuc8; Loening et al., 2006, 2007) was fused in frame at the C-terminus of the sequence of triple HA-tagged (N-terminus) human β2AR (GenBank accession no. AF203386.1; Missouri S&T cDNA Resource Center, Rolla, MO). To generate the resulting P3HB2ALuc-2 plasmid (Gimenez et al., 2012), the sequence of the β2AR was amplified by PCR using the forward primer 5′…gctagaattctgcgatcgcaccaccatggcgtacccatacgatgttcca…3′, which introduces EcoRI and AsiSI restriction sites upstream of the receptor sequence start codon, and the reverse primer 5′…agcggaagcttctagcctgcaggtgccagcagtgagtcatttg...3′, which adds a SbfI restriction site upstream of the receptor coding sequence. The amplified sequence was subcloned into a pCDNA3-based plasmid that included the Rluc8 sequence with a linker with the EcoRI, AsiSI, and SbfI sites.
BRET Experiments.
Arrestin recruitment after agonist stimulation was assessed by BRET-based assays. To characterize the binding profile of β-arrestin 2 to β2AR, COS-7 cells (American Type Culture Collection, Manassas, VA) seeded onto 60-mm cell culture dishes (1.5 × 106 cells/dish) were transiently transfected with a fixed amount of β2AR-RLuc construct (0.5 µg) and varying amounts of Venus arrestin (0–12 µg) using Lipofectamine 2000 (Life Technologies). Transfected levels of β2AR-RLuc are roughly equivalent to endogenous levels (Gimenez et al., 2012). Furthermore, it was shown that manipulation of very few receptor discriminator residues greatly enhances receptor specificity for arrestins, indicating that there is unlikely significant competition of expression vectors with endogenous receptors that are at about the same level in COS-7 cells (Mundell et al., 1999). Maximum arrestin expression in our BRET studies was estimated to be 10-fold over endogenous levels. The following day, cells were transferred to white 96-well plates (5 × 104 cells/well). Luminescence at 460 and 535 nm was measured 48 hours post-transfection after incubating 5 µM coelenterazine-h and the indicated amount of adrenoceptor ligand (or the corresponding vehicle for unstimulated cells) for 5–35 minutes. Net BRET after drug treatment was plotted as a function of the acceptor (Venus arrestin) fluorescence over donor (receptor-RLuc) luminescence (F/L) ratio. Acceptor (Venus arrestin) fluorescence was determined from cells plated onto a black 96-well plate (excitation/emission: 485/535 nm). F/L ratios were used as an indirect measure of arrestin to receptor expression. Arrestin recruitment data (mean ± S.E.M.; eight replicates per experiment) were fit by nonlinear regression to a three-parameter, variable slope hyperbola equation (varying arrestin expression saturation curves) using Prism 6.05 software (GraphPad Software, San Diego, CA). Maximum BRET signal (BRETMAX) was obtained from the curve fits. BRET signals corresponding to different F/L ratios were obtained by interpolating the indicated ratio (e.g., 0.25 or 1) into the resulting curves from the fitted data.
Intact Cell cAMP Measurement.
Human airway smooth muscle (HASM) cells (6–10 passages; kindly provided by Dr. Robert Moore, Baylor College of Medicine, Houston, TX), were grown to confluence in Ham’s F-12 medium in 12-well plates. Although β2AR levels were not routinely measured, they were at the lower limit of detection by [125I]iodocyanopindolol binding, and were in the range of 10–30 fmol/mg. Before experiments, the growth medium was removed and replaced with 0.3 ml NaHCO3-free, 25 mM HEPES-buffered medium (pH 7.4) containing 12 µCi [2,8-3H]adenine at 37°C. For all experiments, the adenine prelabeling time was 3 hours. For protocols including PDE inhibition, 1.0 mM 3-isobutyl-1-methylxanthine (IBMX) was added 30 minutes before agonist treatment. For measuring the time course of agonist stimulation of cAMP, cells were stimulated with either vehicle (AT 1:100, or DMSO 0.5%, for controls), 1.0 μM isoproterenol, or 50 nM salmeterol for the indicated times. To stop the reaction, incubation medium was removed, solution containing 5% trichloroacetic acid, 1.0 mM ATP, and 0.1 mM cAMP was added, and cAMP was purified by a two-stage column (Dowex 50/alumina; Bio-Rad Laboratories, Hercules, CA) procedure (Salomon et al., 1974; Baameur et al., 2014).
cAMP accumulation was also measured by immunoassay; however, this assay was not as sensitive (higher baseline) as the [3H]adenine assay, similar to that reported previously (Rosethorne et al., 2010). Otherwise shaping of the cAMP accumulation was identical for both assays. [3H]adenine uptake into the ATP pool is completed by 2 hours and remains stable thereafter for prolonged agonist treatment times. The level of ATP labeling by [3H]adenine is not diminished by 12-hour salmeterol pretreatment.
Two-Step Isoproterenol Rescue Protocol.
For measurement of the effects of salmeterol pretreatment on subsequent isoproterenol stimulation of cAMP (rescue), cells were pretreated with 20 nM salmeterol for either 10 minutes, 2 hours, or 14 hours. Before the experiments, cells were prelabeled with [2,8-3H]adenine with or without the addition of IBMX as described above. After salmeterol pretreatments, cells were stimulated with 100 μM isoproterenol (with or without 1.0 mM IBMX) for 0–30 minutes and cAMP was measured. All assay points were performed in triplicate at each time of treatment. The average values of triplicates for multiple experiments were expressed as the mean ± S.E.M.
Results
Comparison of Salmeterol- and Isoproterenol-Liganded β2AR Interactions with β-Arrestin 2 by BRET.
Prior biochemical and modeling studies demonstrated reduced salmeterol-mediated translocation and internalization of the β2AR relative to isoproterenol. Based on this, we proposed that the affinity of the salmeterol-β2AR complex for arrestin was much reduced (Vayttaden et al., 2010). To address this proposal, we initiated a BRET study to determine the effects of ligands on the recruitment of arrestin to the liganded β2AR using transient expression of Venus N-terminally tagged β-arrestin 2 and the β2AR-RLuc8 fusion proteins. In each experiment, arrestin levels (directly proportional to fluorescence) were varied at a constant β2AR-Rluc (luminescence) expression level, giving six F/L ratios that varied from 0.25 to 3.0 (Fig. 1, A–D). BRET was assessed from 5 to 35 minutes after the addition of either 100 nM salmeterol, 10 μM isoproterenol, or the inverse agonists ICI-118.551 (1.0 μM) and carazolol (0.1 μM). Fig. 1D shows a comparison of the effects of challenges with high β2AR occupancy by these compounds on BRETMAX (F/L = 3.0) signals. The data showed a much reduced BRETMAX (F/L = 3.0) with salmeterol relative to isoproterenol. Furthermore, although BRET with isoproterenol steadily diminished over 10–35 minutes, the salmeterol signal was relatively constant. ICI-118.551 and carazolol initially showed diminished BRET relative to the control, which in time approached baseline, consistent with their inverse agonism.

Arrestin recruitment by salmeterol is much reduced compared with full agonists such as isoproterenol. Maximum arrestin recruitment (BRETMAX) by ligand-occupied β2ARs was determined by type 1 BRET-based assays. (A–D) Recruitment of various expression levels of N-terminal fused Venus β-arrstin2 by a fixed amount of C-terminal fused β2AR-RLuc after incubation for the indicated time points with 10 μM isoproterenol (open triangles and solid lines) (A), 0.1 μM salmeterol (filled circles, solid line) (B), 1 μM ICI-118.551 (ICI, open diamonds, dashed line) (C), and 0.1 μM carazolol (filled squares, dashed line) (D). Data represent the net BRET after subtraction of BRET ratios obtained in the absence of ligand (y-axis) plotted as a function of (x-axis) the F/L ratio. BRETMAX was determined after fitting the data with a three-parameter (variable slope) hyperbolic function. Data points represent the mean ± S.E.M. (n = 8) from a typical experiment. (E) Net BRETMAX (F/L = approximately 3) for the different ligands used in (A) to (D) at the indicated (x-axis) time points. Cara, carazolol; ICI, ICI-118.551; Iso, isoproterenol; Salm, salmeterol.
To investigate the interaction at reduced agonist occupancy, we performed the experiments shown in Fig. 2 with the following treatments: 1) 30 and 100 nM isoproterenol, since previous studies showed that these concentrations were either at or slightly under the Kd for isoproterenol, but produced maximal GRK phosphorylation in time (Tran et al., 2004); and 2) 10 and 30 nM salmeterol, which exceeded the Kd (1 to 2 nM) but were used to compensate for the much diminished BRET signal. The results from the data shown at three F/L ratios (3.0, 1.0, and 0.25) demonstrated first that salmeterol at either concentration caused a very weak but significant signal that increased slightly in time, with BRETMAX similar to that with 100 nM salmeterol (Fig. 1D). Second, isoproterenol at either 30 or 100 nM caused a BRET signal that diminished significantly over 10–35 minutes. BRETMAX with 100 nM isoproterenol was equivalent to that found with 10 μM isoproterenol (Fig. 1); at 30 nM, the signal was significantly decreased in agreement with the GRK phosphorylation profile (Tran et al., 2004).

Reduction in salmeterol-induced arrestin recruitment is independent of the arrestin to receptor (F/L) expression level. β-arrestin 2 recruitment was assessed by BRET at three different arrestin to receptor expression-level (F/L) ratios: F/L = approximately 3 (BRETMAX) (A), F/L = 1 (B), and F/L = 0.25 (C). The assays were performed in the presence of 10 nM (filled circles, dashed lines) and 30 nM (open circles, dashed lines) salmeterol, or 30 nM (filled triangles, solid line) and 100 nM (open triangles, solid line) isoproterenol. Experiments were performed as in Fig. 1, and net BRET values at the different F/L ratios were obtained after interpolating the BRET values for the respective F/L ratios after fitting the data to three-parameter variable slope hyperbolas (raw data not shown). Values represent the mean ± S.E.M. (n = 8 replicates, two independent experiments). Iso, isoproterenol; Salm, salmeterol.
To assess the effect of varied arrestin levels and the time of assay, we calculated the ratios of BRET signals (BRETISO/BRETSALM) from the data of Fig. 2. The results shown in Fig. 3 demonstrate a striking effect of the time of assay of the arrestin/β2AR-ligand complex at all three F/L levels. With F/L ratios of 3.0 and 1.0, the isoproterenol-mediated signal at 5 minutes was approximately 20-fold higher than for salmeterol. For the F/L ratio of 0.25, the isoproterenol/salmeterol signal was approximately 35, likely reflecting greater error caused by the reduced BRET signal at low arrestin levels.

Arrestin recruitment by isoproterenol significantly diminishes over time in contrast with salmeterol’s weaker albeit persistent effect. To highlight the differences in the arrestin recruitment time course (5–35 minutes) between isoproterenol (Iso; 100 nM) and salmeterol (Salm; 30 nM), the BRET data from Fig. 2 were transformed by calculating the ratios (Iso/Salm) between the net BRET values for F/L = approximately 3 (BRETMAX) (A), F/L = 1 (B), and F/L = 0.25 (C) for each drug, respectively. Values represent the mean ratio ± range (highest to lowest) from two independent experiments. Iso, isoproterenol; Salm, salmeterol.
The time dependence of the BRET results for isoproterenol agree well with the kinetics of its rapid stimulation of GRK phosphorylation, arrestin binding, and internalization of the β2AR (t1/2 of internalization = approximately 3 minutes); upon internalization, isoproterenol and arrestin dissociate from the receptor, consistent with the transient nature of the arrestin-receptor complex under these circumstances (Tran et al., 2004; Vayttaden et al., 2010). By contrast, salmeterol-induced BRET increased slightly in time (5–10 minutes) and stabilized, demonstrating salmeterol’s reduced efficacy for recruitment of the arrestin. These findings show that although low efficacy limits the level of the salmeterol-β2AR-arrestin complex, the stability of the complex is sufficient to cause limited internalization. The results are consistent with the previously reported very limited salmeterol-induced translocation and internalization data (Moore et al., 2007), as well as with the simulations of the GRK pathway for isoproterenol versus salmeterol (Vayttaden et al., 2010).
The Role of PDE in Isoproterenol and Salmeterol Stimulation of cAMP Accumulation and Functional Desensitization.
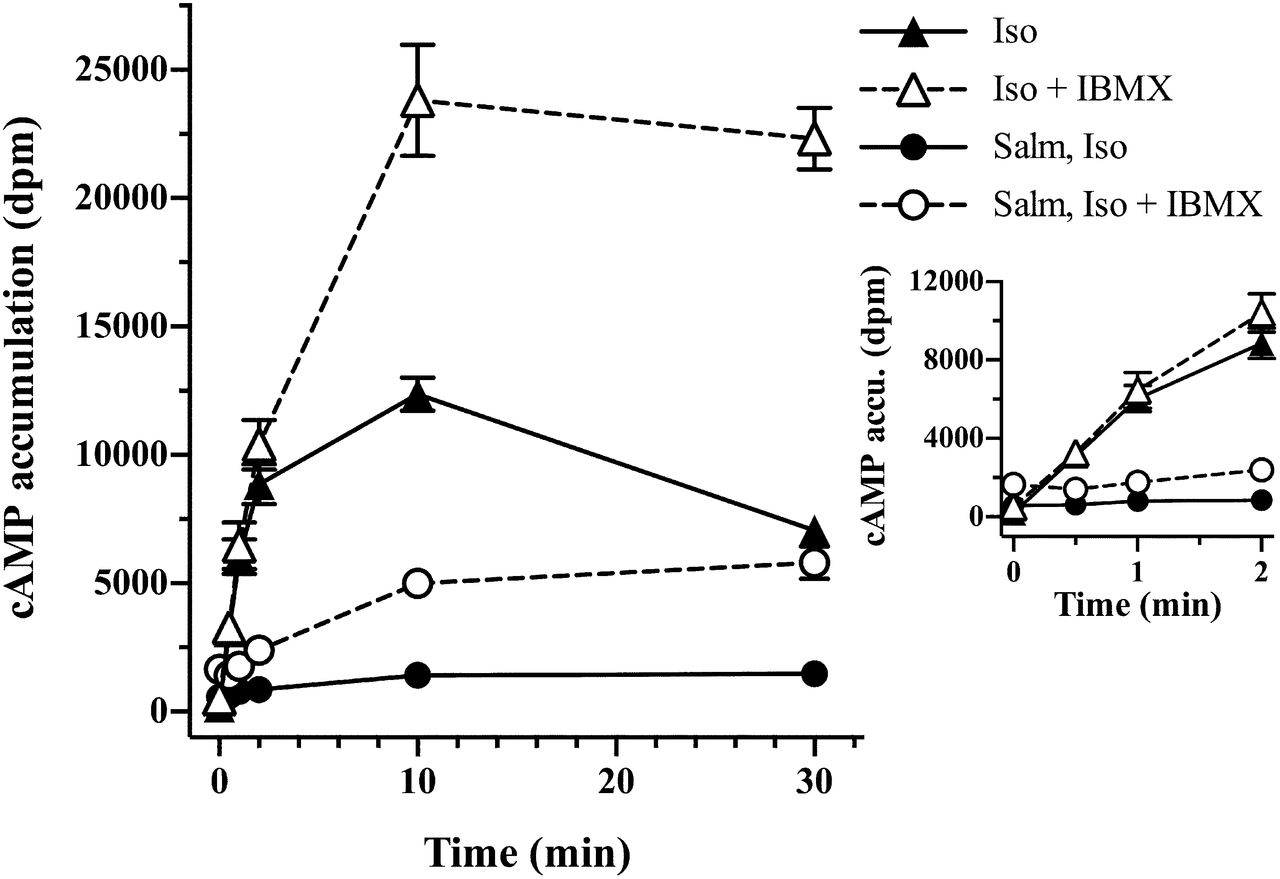
The goals of this aspect of our study were to examine and contrast isoproterenol and salmeterol stimulation of cAMP turnover in primary cultured HASM cells, to determine the effects of salmeterol pretreatment on subsequent isoproterenol stimulation, and to allow comparisons of the extent of functional desensitization (cAMP levels) with and without PDE inhibition. To that end, we first evaluated the effect of the presence and absence of IBMX (1.0 mM) on cAMP accumulation in HASM cells in response to agonist, as shown in Fig. 4A. We found that the efficacy of salmeterol stimulation in the presence or absence of PDE inhibition was much reduced relative to isoproterenol and formoterol over the entire time course, consistent with previous findings with low endogenous levels of the β2AR (Clark et al., 1996; Rosethorne et al., 2010). All agonists demonstrated functional desensitization shown by the rapidly diminishing rates of cAMP accumulation in time (without IBMX), with cAMP levels peaking at 5 minutes and subsequently declining. In the presence of IBMX to moderate the confounding effects of PDE (added 30 minutes before agonists), cAMP levels at time zero were slightly augmented relative to controls, showing the effect of the pretreatment with IBMX. With agonist stimulation, IBMX eliminated the rapid rise and fall of cAMP and levels increased to plateau values. The progressive increase in cAMP levels caused by IBMX demonstrates the time dependence of the downstream functional desensitization by PDE, although inhibition of PDE was not complete (see Discussion below). Nonetheless, the shaping of the cAMP levels reflects primarily receptor-level desensitization by all agonists.

Time course of cAMP accumulation in HASM cells after stimulation with isoproterenol, formoterol, or salmeterol in the presence and absence of 1.0 mM IBMX. (A) HASM cells were prelabeled with [3H]adenine as described in the Materials and Methods and preincubated with or without 1.0 mM IBMX for 30 minutes, after which either isoproterenol (1.0 μM), formoterol (100 nM), or salmeterol (50 nM) was added. cAMP values were from one experiment (representative of n = 2 for formoterol, n = 4 for salmeterol, and n = 6 for isoproterenol). (B) Initial rates of cAMP formation (0–2 minutes) were followed in HASM cells treated a described above with either salmeterol or isoproterenol (with or without IBMX) as indicated. Values are the mean ± S.E.M. (n = 3). Form, formoterol; Iso, isoproterenol; Salm, salmeterol.
To more clearly compare initial rates (0–2 minutes) of cAMP accumulation with isoproterenol and salmeterol with or without IBMX, a separate series of experiments was performed (Fig. 4B). Although the low levels of stimulation (without IBMX) generate significant noise, it was clear that the efficacy for salmeterol is reduced approximately 80% relative to isoproterenol (note values at 1 and 2 minutes). In the presence of IBMX, the initial rates were unmodified; however, after 2-minute stimulation, there was expectedly a stimulatory effect of PDE inhibition with isoproterenol as levels of cAMP approached levels such that PDE hydrolysis was significant relative to rates of agonist activation.
In preliminary studies, we found that 1.0 mM IBMX, 50 mM rolipram, or their combination resulted in similar levels of cAMP augmentation, and maximum effects of IBMX were achieved at 250 µM (data not shown). We also examined the extent of PDE inhibition by observation of the rates of decay of cAMP after 10-minute isoproterenol stimulation with addition of propranolol. The rates we observed (t1/2 = 3.98 ± 0.68 minutes and 1.85 ± 0.28 with and without propranolol, respectively; n = 4) showed 54% inhibition. Given the affinity of IBMX for PDE, full inhibition of PDE activity is theoretically unattainable, as previously discussed (Xin et al., 2008). However, it is important to realize that removal of isoproterenol activation will diminish PKA activation of PDE in time, and it is likely that its inhibition is greater than that measured by following the decay of cAMP.
The important conclusion from the agonist shaping of cAMP levels is that salmeterol causes an apparent desensitization that approaches that of isoproterenol and formoterol. A more quantitative estimate of their relative extents of functional desensitization after 15 minutes was performed by comparison of the initial and final rates of cAMP accumulation in the presence of IBMX (Fig. 4). At any point in time, the following equation describes cAMP levels: where kact is the rate of cAMP formation, and kd is the rate of hydrolysis of cAMP at time t.
where kact is the rate of cAMP formation, and kd is the rate of hydrolysis of cAMP at time t.
First, Fig. 4 shows that the initial rate of salmeterol stimulation was just 20% that of the strong agonists. Second, we can compare the relative activities of agonists at the point at which d[cAMP]/dt is approximately = 0, where kact = kd [cAMP]. If we assume that at the plateau values of cAMP accumulation, the rate of hydrolysis by PDE (kd) is equivalent for all agonists (Nino et al., 2009), then we can compare kact for the two agonists. Substituting the cAMP values at 15 minutes (plus IBMX conditions) for salmeterol (approximately 5000) and isoproterenol (approximately 15,000) into the equation kact = kd [cAMP] for each agonist demonstrates that kact for isoproterenol is approximately 3-fold greater than for salmeterol, whereas it was 5-fold greater based on initial rates. Thus, the relatively greater loss of isoproterenol activity reflects greater desensitization. An important further consideration is the extent to which spare receptors in HASM cells affect desensitization by strong versus weak agonists. The effects of spare receptors in a variety of non-HASM cells were well characterized previously (Whaley et al., 1994; January et al., 1998; Clark and Knoll, 2002; Xin et al., 2008). Using human bronchial preparations and measurement of bronchodilation, it was recently shown that they have sufficient receptor reserve (Giembycz, 2009) such that for the strong agonist formoterol, the EC50 for bronchodilation was left-shifted approximately 120-fold from the Kd and to a much greater extent than for weak agonists. Although our primary cultures of HASM cells likely have much lower levels (10–30 fmol/mg), we have found that the EC50 for isoproterenol is approximately 8-fold left-shifted relative to the Kd (EC50 = 30 nM; Kd for isoproterenol = 250 nM). The result is that salmeterol desensitization will cause immediate decreases in Emax, whereas isoproterenol will likely initially display increases in EC50 with limited change in Emax, thus further overestimating the desensitization caused by salmeterol relative to isoproterenol. The most parsimonious explanation for the reduced salmeterol functional desensitization consistent with the BRET result is the much reduced salmeterol recruitment of arrestin, which in turn causes reduced internalization, even while GRK phosphorylation eventually reaches a level (10–15 minutes) similar to isoproterenol (Tran et al., 2004).
Effect of Pretreatment with Salmeterol on Subsequent Stimulation by Isoproterenol.
As discussed in the Introduction, studies of salmeterol-mediated desensitization are confounded by its rapid partition into membranes, precluding washout studies with subsequent restimulation (Rhodes et al., 1992; Clark et al., 1996; January et al., 1998; Lombardi et al., 2009). To overcome this obstacle, we used a two-stage protocol in which HASM cells were first stimulated with 20 nM salmeterol for either 10 minutes (Tran et al., 2004), 2 hours, or 14 hours after which a high concentration of isoproterenol (100 µM) was added to overcome salmeterol stimulation, importantly replacing the need for washout. All assays were performed with 1.0 mM IBMX added 30 minutes before isoproterenol. Since the Kd for salmeterol is 1 to 2 nM and that for isoproterenol is approximately 200 nM, occupancy by isoproterenol (500-fold over the Kd) is sufficient to displace salmeterol. The results shown in Fig. 5 for prestimulation by salmeterol for either 10 minutes (squares) or 2 hours (closed triangles), or isoproterenol and salmeterol with no prestimulation as controls, demonstrated several important features. First, the effect of isoproterenol stimulation after each of the salmeterol pretreatment times showed a slight lag (1 minute) in initial rates of isoproterenol. After the lag, isoproterenol caused profound further stimulation, although it was much reduced compared with the control isoproterenol stimulation alone, with the inhibitory effect of a 2-hour pretreatment greater than that after 10 minutes of pretreatment. The lag precluded estimates of desensitization from initial rates. This lag may be attributed both to the slow off rate of salmeterol binding as well as a memory of GRK phosphorylation by salmeterol “priming the system” for subsequent isoproterenol stimulation of arrestin binding (Vayttaden et al., 2010). A second feature of the shaping of the cAMP accumulation is that isoproterenol produced further desensitization after salmeterol prestimulation, indicating that salmeterol-induced functional desensitization of the isoproterenol response (approximately 30–50% at the 10-minute points) was not as extensive as that subsequently caused by isoproterenol even after 2 hours of salmeterol. cAMP levels also approached the control isoproterenol stimulation after 30 minutes with salmeterol pretreatment, a further indication of the lesser desensitization caused by salmeterol relative to isoproterenol. As a further control, costimulation by 20 nM salmeterol and 100 μM isoproterenol with no pretreatment (data not shown) was not significantly different from that of isoproterenol alone.

Effect of a 10-minute and 2-hour preincubation with 20 nM salmeterol on the time course of isoproterenol stimulation of cAMP accumulation. HASM cells prelabeled with [3H]adenine as described in the Materials and Methods were pretreated with 20 nM salmeterol for either 10 minutes or 2 hours, and 1.0 mM IBMX was added to all incubations for 30 minutes before the assay of cAMP. After preincubation of salmeterol, cells were stimulated with 100 μM isoproterenol and cAMP accumulation followed for 2–30 minutes. Values are the means ± S.E.M. of triplicate determinations and are representative of two independent experiments. Iso, isoproterenol; Salm, salmeterol.
Effect of a 14-Hour Pretreatment with Salmeterol on Isoproterenol Stimulation of cAMP Accumulation.
The salient clinical efficacy of salmeterol is derived from its prolonged bronchodilation (10–12 hours). To assess characteristics of its long action, we examined the effects of a 14-hour pretreatment with salmeterol on subsequent isoproterenol stimulation of cAMP accumulation in HASM cells. Cells were pretreated with or without 20 nM salmeterol for 14 hours and were then stimulated with 100 μM isoproterenol for 0–30 minutes in the presence or absence of IBMX (Fig. 6). This allowed observation of the salmeterol-mediated desensitization of the initial rates of isoproterenol stimulation (Fig. 6, inset), as well as the subsequent further desensitization induced by isoproterenol relative to controls. In the absence of IBMX (closed symbols), salmeterol pretreatment caused a 3-fold increase in basal values (time zero). After 2 minutes, isoproterenol-stimulated levels were not significantly elevated over basal values. After 10–30 minutes, isoproterenol-stimulated levels were 3-fold over basal. Although the extent of desensitization appeared to be >95% based on the initial rate of cAMP accumulation relative to isoproterenol stimulation in controls (no salmeterol), it rather likely reflects the slow off rate of salmeterol. Therefore, to estimate functional desensitization, we compared the fold activation of isoproterenol over basal values at the 10-minute points (3-fold with salmeterol pretreatment versus approximately 40-fold without), which indicated >90% salmeterol-induced desensitization. However, this overestimates the desensitization since the salmeterol pretreatment increases the basal cAMP value.

Effect of a 14-hour preincubation with 20 nM salmeterol on the time course of cAMP accumulation. HASM cells were pretreated with or without 20 nM salmeterol for 18 hours, after which cells were incubated with or without 1.0 mM IBMX for 30 minutes before the addition of 100 μM isoproterenol. cAMP accumulation was followed from 0.5 to 30 minutes. Values are the means ± S.E.M. of three independent experiments. The inset shows the time course of cAMP accumulation from 0.5 to 2.0 minutes. accu., accumulation; Iso, isoproterenol; Salm, salmeterol.
In the presence of IBMX, the basal levels (time zero) were further increased after salmeterol pretreatment, followed by an insignificant elevation of cAMP by isoproterenol over 0–2 minutes (Fig. 6, inset). After 10-minute isoproterenol stimulation, cAMP levels with and without salmeterol pretreatment were approximately 3.5-fold and 100-fold over basal, respectively, indicating that the salmeterol-induced receptor-level desensitization was extensive (approximately 95% after 10- to 30-minute stimulation). Importantly, isoproterenol stimulated additional desensitization of the β2AR, consistent with the conclusion that as extensive as salmeterol desensitization was from the pretreatment, it was not complete relative to isoproterenol. Furthermore, cAMP levels with the salmeterol pretreatment (with IBMX) were in fact 20–30% of those without pretreatment at the 10- to 30-minute time points. By these measures, the apparent salmeterol-induced desensitization of isoproterenol cAMP accumulation was 70–80%. Regardless of the methods for estimating functional desensitization, it was significantly greater than that from either the 10-minute or the 2-hour pretreatments shown in Fig. 5.
Discussion
Salmeterol used in combination with steroids has been very effective in the treatment of asthma and chronic obstructive pulmonary disease. Providing an explanation for the clinical and molecular efficacy of salmeterol has been challenging because of its unique chemical properties and the complexity of agonist-induced desensitization and downregulation of the β2AR. Early studies clearly demonstrated that salmeterol was a weak partial agonist for β2AR activation of AC, and it caused less receptor-level desensitization relative to strong agonists attributable to weak efficacy for stimulation of GRK phosphorylation and sequelae. On the basis of studies of functional desensitization using equieffective concentrations of salmeterol and isoproterenol, it was shown that salmeterol exhibited properties of a strong agonist. In this study, we resolve these paradoxical results by both providing further molecular evidence for salmeterol’s diminished efficacy for stimulation of the GRK pathway, and yet demonstrating through careful analysis of cAMP turnover and the downstream PKA regulation of PDE that salmeterol actions on the G protein–dependent/PKA pathway at high occupancy show strong efficacy.
Considering first the GRK pathway, Vayttaden et al. (2010) modeled and simulated the short-term isoproterenol and salmeterol-stimulated GRK/arrestin/internalization pathway of desensitization in HEK293 cells. We concluded that at least for short-term stimulation, salmeterol exhibits low efficacy for this pathway. A key assumption in the modeling was that the affinity of the salmeterol-β2AR complex for arrestin was greatly reduced relative to isoproterenol. In this study, this assumption was addressed through the use of BRET. We report that salmeterol shows much reduced recruitment of arrestin to the receptor, consistent with our proposal. At the earliest time measured (5 minutes), isoproterenol-stimulated BRET was 20- to 35-fold greater than salmeterol (Fig. 3) at F/L ratios of 3.0 and 0.25, respectively. Between 15 and 20 minutes after stimulation, the ratio stabilized, with isoproterenol-induced BRET approximately 5-fold elevated over salmeterol. Thus, from BRET, the affinity of the salmeterol-β2AR complex for arrestin is in the range of 5–20% that of isoproterenol. Our prior simulations showed that the best agreement to experimental data required a 100-fold reduction in arrestin affinity (Vayttaden et al., 2010).
The decrease in BRET for isoproterenol reflects the rapid isoproterenol-induced internalization and rapid dissociation of isoproterenol and arrestin from the complex (Fig. 7). Over the same time, the salmeterol-β2AR-arrestin complex is much reduced but stable, but sufficient to promote a low level of salmeterol-induced internalization. We previously reported that overexpression of arrestin increased salmeterol-induced arrestin translocation and internalization (Moore et al., 2007). Maximum arrestin expression in our BRET studies was estimated to be 10-fold over endogenous levels, likely contributing further to salmeterol-induced internalization. As previously shown, the initial rate of salmeterol stimulation of GRK phosphorylation was 35% that of isoproterenol; however, the extent of salmeterol stimulation of phosphorylation matched that of isoproterenol after 10–15 minutes (Tran et al., 2004; Moore et al., 2007). Thus, in time, the level of the salmeterol-β2AR-arrestin complex is the rate-limiting step for internalization and desensitization. Interestingly, salmeterol, with about 200-fold higher affinity relative to isoproterenol coupled with its membrane sequestration (as a source of leaked salmeterol), likely stays bound to the small fraction of the receptor after internalization, perhaps further explaining the stabilization of the complex with time (Fig. 7). That is, the BRET signal for salmeterol (in contrast with isoproterenol) would not distinguish between surface and internalized receptors. In summary, our BRET findings strongly support the conclusion that short-term salmeterol desensitization via the GRK pathway is diminished by its reduced efficacy for arrestin binding. In support of this conclusion, the observation of a deficit in salmeterol-induced internalization via GRK/arrestin was also demonstrated in HAMC cells by immunolocalization of yellow fluorescent protein–tagged β2AR after salmeterol stimulation (Cooper et al., 2011), in mouse myoblast C2C12 cells using a galactosidase complementation assay (Carter and Hill, 2005), and in HEK 293 cells by arrestin translocation (van der Westhuizen et al., 2014) and fluorescence resonance energy transfer (Drake et al., 2008). Furthermore, it was also shown that knockdown of arrestin reduces functional desensitization (Deshpande et al., 2008).
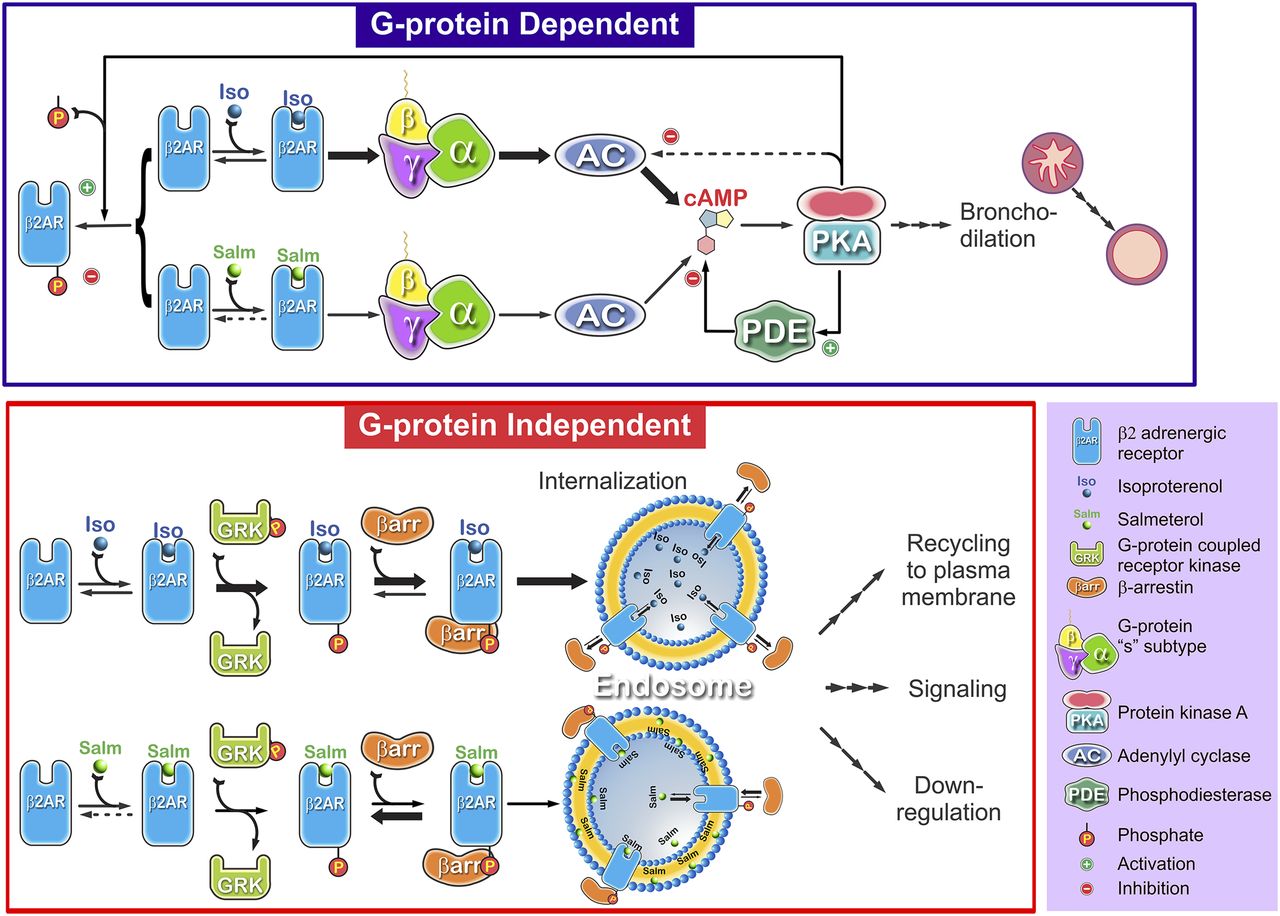
Consequence of reduced salmeterol efficacy in G protein–dependent and G protein–independent pathways of β2AR desensitization. It is well established that strong agonist (isoproterenol/epinephrine)–induced desensitization of the β2AR involves two discrete pathways. The highly amplified Gs-AC-PKA (G protein–dependent) pathway leads to PKA phosphorylation of the β2AR and partial uncoupling, PDE activation, and ultimately bronchodilation (possible PKA phosphorylation and inhibition of AC subtypes is not shown). The occupancy-dependent G protein–independent pathway (lower panel) is initiated by ligand-bound β2AR activation of GRK phosphorylation. GRK phosphorylation leads to β-arrestin binding to the ligand-bound, GRK-phosphorylated β2AR and complete uncoupling of Gs activation. Arrestin binding triggers clathrin-coated pit internalization. Once internalized, the low-affinity agonists (e.g., isoproterenol) and arrestin dissociate. Because of its stabilization of a unique state of the β2AR, salmeterol shows reduced efficacy for activation of AC (at endogenous β2AR levels), causing reduced accumulation of cAMP relative to isoproterenol (thick versus thin arrows). However, because of the amplification caused by the positively cooperative cAMP activation of PKA, it causes similar levels of PKA-modulated actions at high occupancy relative to strong agonists. For the GRK pathway, salmeterol-β2AR exhibits reduced efficacy for GRK activation and arrestin binding. The low level of the salmeterol-β2AR-arrestin complex drives much reduced internalization and resultant levels of endosomal β2AR. In this diagram, we indicate another possible difference in salmeterol versus isoproterenol; that is, binding and rebinding occurs because of the extreme partitioning of salmeterol into the plasma membrane coupled with its high affinity for receptor, prolonging β-arrestin binding.
In the second aspect of these studies, we attempted to parse out the relative contributions of receptor-level desensitization and downstream PDE activity to β2AR desensitization observed in HASM cells. This was accomplished by two approaches: 1) direct observation of the rapid kinetics of cAMP turnover, and 2) a simple two-stage rescue protocol, in which cells were first stimulated with salmeterol and then challenged by a second, high-occupancy stimulation with isoproterenol to counter the inability to washout sequestered salmeterol. With regard to the kinetics of salmeterol stimulation of cAMP turnover in the absence of IBMX, the rapid decline in the rate of cAMP accumulation after peak levels demonstrated significant β2AR desensitization. Inclusion of IBMX demonstrated the large effects of PDE inhibition (approximately 70–80% augmentation of cAMP at the 15-minute time points) and hence the major role of PKA phosphorylation of PDE in desensitization. Estimation of the efficacy for salmeterol-mediated desensitization with much reduced contribution from PDE activity was accomplished by analysis of the shaping of cAMP accumulation in the presence of IBMX by all agonists (Fig. 4). Several observations indicated that salmeterol desensitization was reduced relative to isoproterenol. First, calculations of the initial versus final rates of cAMP accumulation indicated less salmeterol desensitization. Second, we used high occupancy of agonists, and this in conjunction with significant but limited receptor reserve in HASM cells relative to that of human bronchial preparations (Giembycz, 2009) predicts that the reduction in cAMP levels Emax by salmeterol relative to isoproterenol are likely overestimated. That is, with receptor reserve, the effects of strong agonists on concentration-response curves after stimulation are to increase EC50 until receptor levels are much reduced, whereas salmeterol will cause decreases in Emax (Whaley et al., 1994; Clark et al., 1999). From simulations of salmeterol desensitization via the GRK pathway, we estimated that β2AR activity remaining with either a 100-fold, 10-fold, or no reduction in arrestin affinity for the salmeterol-β2AR complex was 65, 35, and 20%, respectively, after 30 minutes (Vayttaden et al., 2010). Prior biochemical evidence of salmeterol desensitization (effects on cell-free AC stimulation) showed it was just 20% of that caused by high-efficacy agonists over the short term in several cell systems (Clark et al., 1996; January et al., 1998; Vayttaden et al., 2010). In summary, the kinetics of cAMP turnover combined with the BRET findings demonstrate that salmeterol causes much reduced desensitization attributable to the GRK pathway, but salmeterol nonetheless causes substantial desensitization by acting as a strong agonist in the cAMP/PKA pathway.
The rescue protocol demonstrated that salmeterol-induced desensitization, estimated by cAMP levels in the presence of IBMX at the 10-minute time points, progressed from a moderate extent after 10-minute or 2-hour stimulation (approximately 30–50%) to 75–80% after 14 hours. Furthermore, the data indicate that even with the 14-hour pretreatment, isoproterenol was still able to stimulate the receptor after the lag and cause further desensitization, as indicated by the turnover of cAMP reaching a plateau value between 10 and 30 minutes. Since downstream PKA activation of PDE is both rapid and prolonged and is accompanied by PDE induction as well (Xin et al., 2008; Nino et al., 2009, 2012), it is likely that the contribution of PDE was relatively constant over the time of isoproterenol stimulation, regardless of the time of salmeterol pretreatment. This is further supported by the similar IBMX-induced increase in cAMP levels after isoproterenol (10–30 minutes). In addition, receptor phosphorylation by PKA should be equivalent for salmeterol and isoproterenol because it is occupancy independent. To summarize our findings with the rescue protocol, it is clear that although salmeterol caused significant and progressive desensitization of isoproterenol stimulation with pretreatments, further desensitization by isoproterenol occurred. The most parsimonious explanation for the reduced salmeterol desensitization relative to isoproterenol is its low efficacy for activation and arrestin binding. In addition, salmeterol prestimulation primes the receptor by the “memory” of salmeterol-stimulated GRK phosphorylation, such that addition of isoproterenol leads to rapid arrestin binding (Vayttaden et al., 2010).
Since salmeterol-mediated desensitization is caused by both PKA and GRK phosphorylation of the β2AR and PDE activation by cAMP/PKA, it is not surprising that there has been ambiguity in defining the overall efficacy for salmeterol functional desensitization. It has also been suggested that there may be a 30–35% reduction in AC activity (types 5 and 6) from PKA phosphorylation in HASM cells, further contributing to PKA-regulated events (Xu et al., 2001; Horvat et al., 2012). Therefore, our findings using high occupancy by agonists are actually consistent with the results from several groups, in which equieffective concentrations of agonists were used to assess the functional end point (i.e., desensitization of bronchodilation, and bronchoprotection caused by long-term treatments with salmeterol relative to strong by agonists; Szczuka et al., 2009; Cooper et al., 2011). Equieffective concentrations means that comparisons were made using very low occupancy of strong agonists with high-occupancy salmeterol to achieve similar cAMP stimulated levels. Under these conditions, salmeterol-induced functional desensitization would be expected to be approximately equivalent to that of isoproterenol or formoterol, since the occupancy-dependent GRK pathway for strong agonists would be minimized.
Another important aspect in the assessment of salmeterol’s long-term effects is the very slow downregulation process. Interestingly, we previously reported that dobutamine, a very low efficacy agonist for AC stimulation and GRK phosphorylation, showed a level of downregulation in the normal human lung epithelial cell line, BEAS-2B, equivalent to isoproterenol (Williams et al., 2000). Although some inroads have been made to determine the mechanism of the shunting of the β2AR either to the recycling pathway or degradation (Liang et al., 2008), salmeterol-mediated downregulation in HASM cells has not been explored, primarily due to the problems of its sequestration in membranes and the limitations of receptor antibodies at endogenous levels of β2AR. Given the fact that downregulation is a very slow process, it is possible that there is a logjam of a key intermediate in downregulation such that weak agonists like dobutamine, and possibly salmeterol, cause downregulation equivalent to epinephrine, acting as a strong agonist in this scenario.
It is interesting to speculate on the implications of our findings to the clinical use of salmeterol with the obvious caveat that primary cultures of HASM cells, while a very useful model, differ quite considerably from in vivo use of the drug. In addition, our studies focused on the GRK pathway and the role of only one downstream action of PKA (PDE), and it is appreciated that the downstream actions caused by β-adrenergic agonists are complex and are not directly correlated with cAMP levels. However, several recent studies have supported the primacy of PKA in the regulation of bronchodilation. In particular, a recent article by Morgan et al. (2014) concluded that PKA is the primary mechanism by which β-agonists relax airway smooth muscle and specifically discounts the role of Epac proteins. However, PKA activity was not directly measured. Horvat et al. (2012) investigated, for the first time, a potential role of membrane localization by AKAPs in HASM cells. Although they provided good evidence for a minor role of AKAPs in prolonging transient signaling, the authors found that AKAP disruption had “minimal effects on whole-cell cAMP accumulation.” Furthermore, Billington et al. (2008) reported that “elevation of cAMP within the cytoplasm after β2AR stimulation is rapid and shows no distinct spatial compartmentalization in HASM cells.” In terms of downstream actions, BinMahfouz et al. (2015) thoroughly investigated a spectrum of important downstream actions of LABAs using BEAS2B cells, particularly the roles of PDE3 and PDE4 inhibitors on the time dependence of PKA activation of cAMP response element-binding protein and glucocorticoid regulation of glucocorticoid response elements. However, the authors did not investigate any of the efficacy and desensitization parameters that we examined in our study; nonetheless, the studies are remarkably complementary, although they used the LABA formoterol and not salmeterol.
As a further consideration, in the rescue protocol we used an overwhelming concentration of isoproterenol to obviate the problem of sequestered salmeterol after pretreatments. In vivo, it is unlikely that salmeterol would be retained at these high levels due to gradual diffusion from its site of activation (hence the 12-hour limit of salmeterol for effective bronchodilation); thus, our protocol may exaggerate desensitization. Similarly, endogenous levels of epinephrine could never be this high. However, in the instance in which high levels of strong agonists are used as rescue therapy in the clinic, our protocol closely mimics what occurs in vivo since levels of salmeterol would be diminished greatly after 12 hours and rescue agonist is used at high concentrations. Furthermore, the use of salmeterol in the clinic is now strictly indicated for use as an add-on to therapy with steroids. It has been appreciated that use of PDE4 inhibitors in the treatment of asthma and chronic obstructive pulmonary disease may be clinically important and synergize with glucocorticoids and LABA treatment to reduce the frequency of exacerbations, as recently discussed (Kaur et al., 2008; Nino et al., 2010; Cooper et al., 2011; Holden et al., 2011; Dekkers et al., 2012; Manetsch et al., 2013; Moodley et al., 2013; Theron et al., 2013; Giembycz and Newton, 2014; BinMahfouz et al., 2015). Our findings showing the dramatic effects of PDE inhibition on cAMP levels after either long-term or short-term salmeterol treatment support the importance of developing multidrug therapy that includes PDE inhibitors, LABAs, and steroids. Finally, because of salmeterol’s low efficacy, it will act as a competitive antagonist of epinephrine binding in vivo and will thereby possibly reduce epinephrine desensitization in vivo.
In summary, our results suggest that the beneficial effects of salmeterol relative to strong agonists for long-term protection in the presence of steroids derive primarily from its reduced efficacy for AC activation as well as the GRK pathway and its remarkable stability, a combination of its high affinity and membrane sequestration. This, coupled with the highly amplified PKA/PDE-mediated desensitization, better explains the significant functional desensitization that we and others observe with salmeterol since salmeterol activation of PKA will be equivalent to strong agonists even with its low efficacy for AC activation, thus maintaining a significant level of bronchodilation. Finally, it important to put these effects in context with the clinical use of salmeterol with steroids as discussed above, and with recent evidence demonstrating that arrestin also mediates downstream G protein–independent, proinflammatory effects in animal models of airway disease (Walker et al., 2003; Walker and DeFea, 2014). Thus, the beneficial effects of salmeterol may derive not only from the combination of salmeterol’s reduced efficacy for activation, GRK phosphorylation, and arrestin binding but also from its attenuation of arrestin-mediated inflammation, providing additional rationale for its clinical efficacy.
Acknowledgments
The authors thank Ms. Jackie Friedman for assistance in performing the cAMP assays and Vsevolod V. Gurevich (Vanderbilt University) for providing the plasmid construct coding bovine β-arrestin 2.
Authorship Contributions
Participated in research design: Gimenez, Baameur, Vayttaden, Clark.
Conducted experiments: Gimenez, Baameur, Vayttaden.
Contributed new reagents or analytic tools: Gimenez.
Performed data analysis: Gimenez, Baameur, Vayttaden, Clark.
Wrote or contributed to the writing of the manuscript: Gimenez, Baameur, Vayttaden, Clark.
Footnotes
- Received November 12, 2014.
- Accepted March 17, 2015.
↵1 Current affiliation: Signaling Systems Unit, Laboratory of Systems Biology, National Institutes of Health National Institute of Allergy and Infectious Diseases, Bethesda, Maryland.
This research was supported by the National Institutes of Health General Medical Sciences [Grant GM31208 (to R.B.C.)].
Abbreviations
- AC
- adenylyl cyclase
- β2AR
- β2 adrenergic receptor
- BRET
- bioluminescence resonance energy transfer
- DMSO
- dimethylsulfoxide
- F/L
- fluorescence/donor luminescence
- GRK
- G protein–coupled receptor kinase
- HASM
- human airway smooth muscle cell
- IBMX
- 3-isobutyl-1-methylxanthine
- ICI-118.551
- (±)-1-[2,3-(dihydro-7-methyl-1H-inden-4-yl)oxy]-3-[(1-methylethyl)amino]-2-butanol hydrochloride
- LABA
- long-acting β-adrenoceptor agonist
- PDE
- phosphodiesterase
- PKA
- protein kinase A
- U.S. Government work not protected by U.S. copyright





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}