


Abstract
Human organic anion transporter 4 (hOAT4) belongs to a superfamily of organic ion transporters that play critical roles in the body disposition of clinically important drugs, including anti-HIV therapeutics, antitumor drugs, antibiotics, antihypertensives, and anti-inflammatories. In this study, we investigated the role of conserved glycine residues in hOAT4 function. We mutagenized each of the six glycine residues (at positions 11, 241, 383, 388, 400, and 466) to serine, and their functional properties were analyzed in COS-7 cells by measuring the uptake of [3H]estrone sulfate. Our results showed that mutants G11S, G383S, G388S, and G466S exhibited transport activities comparable with those of wild-type hOAT4. In contrast, mutants G241S and G400S almost completely lost transport function. We then further characterized Gly-241 and Gly-400 by mutagenizing these residues to amino acids with varying sizes of side chains, including alanine, valine, and leucine. We demonstrated that increasingly larger side chains at positions 241 and 400 increasingly impaired hOAT4 function. Cell-surface biotinylation using an impermeant biotinylating reagent showed that mutations of Gly-241 and Gly-400 interfered with the trafficking of the transporter onto cell surface. Immunofluorescence analysis of mutant-transfected cells confirmed these results. Substitutions of amino acids with large side chains at positions 241 and 400 resulted in decreased Vmax and increased Km. These results suggest that Gly-241 and Gly-400 are important both in targeting the transporter to the plasma membrane and in substrate binding. This is the first identification and characterization of critical amino acid residues in hOAT4 and may provide important insights into the structure-function relationships of the organic ion transporter family.
Organic ion transporters play critical roles in the body disposition of clinically important drugs, including anti-HIV therapeutics, antitumor drugs, antibiotics, antihypertensives, and anti-inflammatories (Dresser et al., 2001; You, 2002; Koepsell et al., 2003). Organic anion transporters (OATs) and organic cation transporters (OCTs) are two major classes of the organic ion transporter family. So far, four members of the OAT class (OAT1, OAT2, OAT3, and OAT4) and three members of the OCT class (OCT1, OCT2, and OCT3) have been cloned from various species. Although both classes have distinct substrate preferences, with OATs for those carrying negative charges and OCTs for those carrying positive charges, they share several common features: similar predicted transmembrane topology; expression in tissues as varied as kidney, liver, intestine, brain, and placenta; and interaction with numerous structurally and pharmacologically diverse compounds. A hydrophobic backbone and a charge are common requirements of OATs and OCTs for their substrates. An examination of the amino acid sequences of organic ion transporters across species from human to Caenorhabditis elegans revealed six glycine residues that are evolutionarily conserved. We hypothesize that these conserved residues may play critical roles in the function of organic ion transporters. In this study, we used human organic anion transporter 4 (hOAT4) as an experimental model to test this hypothesis.
hOAT4 is an organic anion transporter recently cloned and localized to kidney and placenta (Cha et al., 2000; Ugele et al., 2003). Kidney is one of the major organs responsible for the secretion of endogenous metabolites and xenobiotics, including drugs and environmental toxins, from the body. Placenta forms the sole structural barrier between the mother and the developing fetus. It mediates the transfer of nutrients from the mother to the fetus and eliminates metabolic waste products from the fetus. Multiple transporters in kidney and placenta participate in these processes. Functional characterization in Xenopus laevis oocytes and cultured cells revealed that hOAT4 transports steroid sulfates, such as estrone sulfate and dehydroepiandrosterone-3-sulfate (a major circulating steroid secreted by the adrenal cortex), as well as drugs, such as nonsteroidal anti-inflammatory drugs, antitumor drugs, and environmental toxins (Cha et al., 2000; Babu et al., 2002; Khamdang et al., 2002; Takeda et al., 2002; Ugele et al., 2003). Based on its tissue localization and function, it is therefore proposed that hOAT4 may involve the body disposition of these compounds through kidney and placenta. Despite these crucial roles, the structure-function relationships of hOAT4 have not been elucidated. Using combined approaches of site-directed mutagenesis, functional assay, cell-surface biotinylation, and immunofluorescence analysis, we provide in this study the first identification and characterization of critical amino acid residues in hOAT4 that are important for its transport function.
Materials and Methods
Materials. [3H]Estrone sulfate, [3H]dehydroepiandrosterone 3-sulfate (DHEAS), and [14C]para-aminohippurate ([14C]PAH) were purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). Culture media were obtained from Invitrogen (Carlsbad, CA). All other chemicals were purchased from Sigma-Aldrich (St. Louis, MO).
Generation of hOAT4 and hOAT4-myc cDNAs. hOAT4 cDNA (GenBank accession number AB026198) was amplified by PCR from human placenta cDNA library (BD Biosciences Clontech, Palo Alto, CA). The primers for PCR were designed from the published sequences. The pair of primers was 5′-AGCTGCTAGCTCCAAACAGCAGTTAGGTCAGC-3′ (sense) and 5′-AGCTAAGCTTTGACTAAAGGGGCTCCATGC-3′ (antisense). The cDNA was then subcloned into pcDNA 3.1 vector. hOAT4-myc cDNA encodes a fusion protein consisting of full-length hOAT4 with 10 amino acids of the human c-Myc epitope (EQKLISEEDL) at the carboxyl terminus. The myc-tagged hOAT4 cDNA was synthesized by PCR. The sequences were confirmed by the dideoxy chain termination method.
Site-Directed Mutagenesis. Specific amino acid changes were generated using a QuikChange site-directed mutagenesis kit (Stratagene, La Jolla, CA) following the manufacturer's instructions. The sequences of the oligonucleotides used in the mutagenesis procedure are shown in Table 1. The mutants were confirmed by the dideoxy chain termination method.
Oligonucleotides used for mutagenesis
Expression in COS-7 Cells. COS-7 cells were maintained at 37°C and 5% CO2 in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 100 units/ml penicillin, and 100 μg/ml streptomycin. Cells were transfected with plasmid DNA (hOAT4 wild type, hOAT4 mutants, or pcDNA vector) using LipofectAMINE 2000 reagent (Invitrogen) following the manufacturer's instructions. Twenty-four hours after transfection, the transport activities were measured.
Transport Measurements. Uptake of [3H]estrone sulfate was initiated by adding uptake solution (PBS, pH 7, containing 5 mM glucose and 100 nM [3H]estrone sulfate). At the times indicated in the figures, the uptake was stopped by rapidly washing the cells with ice-cold PBS. The cells were then solubilized in 0.2 N NaOH, neutralized in 0.2 N HCl, and aliquoted for liquid scintillation counting. Uptake count was standardized by the amount of protein in each well. Values were mean ± S.E. (n = 3).
Cell-Surface Biotinylation. Cell-surface expression levels of hOAT4 and its mutants were examined using the membrane-impermeant biotinylation reagent, NHS-SS-biotin (Pierce Chemical, Rockford, IL). The transporters were expressed in COS-7 cells in six-well plates using LipofectAMINE 2000 as described above. After 20 h, the medium was removed, and the cells were washed twice with 3 ml of ice-cold PBS, pH 8.0. The plates were kept on ice, and all solutions were kept ice-cold for the rest of the procedure. Each well of cells was incubated with 1 ml of NHS-SS-biotin (0.5 mg/ml in PBS) in two successive 20-min incubations on ice with very gentle shaking. The reagent was freshly prepared for incubation. After biotinylation, each well was briefly rinsed with 3 ml of PBS containing 100 mM glycine, then incubated with the same solution for 20 min on ice to ensure complete quenching of the unreacted NHS-SS-biotin. The cells were then dissolved on ice for 1 h in 400 μl of lysis buffer (10 mM Tris, 150 mM NaCl, 1 mM EDTA, 0.1% SDS, 1% Triton X-100, and protease inhibitors phenylmethylsulfonyl fluoride, 200 μg/ml, and leupeptin, 3 μg/ml, pH 7.4). The unlysed cells were removed by centrifugation at 13,000 rpm at 4°C. Streptavidin-agarose beads (50 μl; Pierce Chemical) were then added to the supernatant to isolate cell membrane protein. hOAT4 was detected in the pool of surface proteins by polyacrylamide gel electrophoresis and immunoblotting using an anti-hOAT4 antibody (Alpha Diagnostic International, Inc., San Antonio, TX).
Electrophoresis and Immunoblotting. Protein samples were loaded (40 μg/lane) on 7.5% SDS-PAGE minigels and electrophoresed using a mini cell (Bio-Rad, Hercules, CA). Proteins were transferred to PVDF membranes in an electroelution cell (Bio-Rad). The blots were blocked for 1 h with 5% nonfat dry milk in PBS-Tween (80 mM Na2HPO4, 20 mM KH2PO4,100 mM NaCl, and 0.05% Tween 20, pH 7.5), washed, and incubated overnight at 4°C with anti-hOAT4 antibody (2 μg/ml; Alpha Diagnostic International, Inc.). The membranes were washed, incubated with goat anti-rabbit IgG conjugated to horseradish peroxidase (1:15000), and signals were detected by SuperSignal West Dura extended duration substrate kit (Pierce Chemical).
Immunofluorescence of Transfected Cells. Seventy-two hours after transfection, COS-7 cells were washed three times in PBS, fixed for 15 min at room temperature in 4% paraformaldehyde in PBS, and rewashed in PBS. The fixed cells were then permeabilized with 0.1% Triton X-100 for 10 min. After that, the cells were incubated for 15 min at room temperature in PBS containing 5% goat serum and then incubated for 1 h in the same medium containing anti-hOAT4 antibody (3 μg/ml) at room temperature. The cells were washed, and bound primary antibodies were detected by reaction with fluorescein isothiocyanate-coupled goat anti-rabbit IgG (Chemicon International, Temecula, CA) diluted 1:200 for 1 h. Cells were thoroughly washed, and the cover glasses were mounted in GEL/MOUNT (Biomeda, Foster City, CA). Samples were visualized with a Zeiss LSM-510 laser scanning microscope (Carl Zeiss Inc., Thornwood, NY).
Statistics. To test the significance of differences between data sets, Student's t test was performed.
Results
Immunoblot Analysis of hOAT4. To our knowledge, there has been no report on the detection of hOAT4 protein by immunoblot analysis using anti-hOAT4 antibody. Therefore, the expression of hOAT4 protein in COS-7 cells was first examined by immunoblot analysis of cell-surface proteins (Fig. 1). Cell-surface proteins were biotinylated with the membrane-impermeant reagent NHS-SS-biotin, isolated with streptavidin-coated beads, and immunoblotted with anti-hOAT4 antibody. Anti-hOAT4 was raised in rabbit against a synthetic peptide corresponding to the 18 amino acids of the COOH terminus of hOAT4. The antibody recognized a product with a molecular mass of ∼83 kDa in hOAT4-transfected cells (Fig. 1a, lane 1) but not in pcDNA vector-transfected cells (Fig. 1a, lane 2; mock control). To further confirm that this product was hOAT4, we engineered an epitope tag (c-myc) at the C terminus of the hOAT4 protein, allowing the detection of hOAT4 protein using anti-myc antibody. Anti-myc antibody recognized a product (Fig. 1a, lane 3) that has the same molecular mass as that detected by hOAT4 antibody. The predicted molecular mass for hOAT4 from its amino acid sequence is ∼60 kDa. The higher molecular mass of 83 kDa observed in this study may represent the glycosylated form of hOAT4. To test this possibility, we treated the cell-surface proteins from hOAT4-transfected cells with PNGase F, an enzyme specifically cleaving N-linked oligosaccharide chains (Fig. 1b). After the treatment, a 60-kDa product was then detected on the immunoblot. These results indicate that hOAT4 is modified by abundant N-glycosylation in COS-7 cells. Importantly, these results demonstrate the feasibility of using anti-hOAT4 antibody for the following structure-function studies.

Immunoblot analysis of hOAT4 expressed at cell surface. Cell-surface proteins were isolated through biotinylation from cells transfected with hOAT4 (lane 1), hOAT4-myc (lane 3), or pcDNA vector (V) (lanes 2 and 4), separated by SDS-PAGE, and transferred to PVDF membrane (a). The blots were then probed with anti-hOAT4 antibody (lanes 1 and 2) or anti-myc antibody (lanes 3 and 4), followed by horseradish peroxidase-labeled secondary antibody. Cell-surface proteins isolated through biotinylation from hOAT4-transfected cells were treated with (lane 2) or without (lane 1) PNGase F, separated by SDS-PAGE, and transferred to PVDF membrane (b). The blots were then probed with anti-hOAT4 antibody, followed by horseradish peroxidase-labeled secondary antibody.
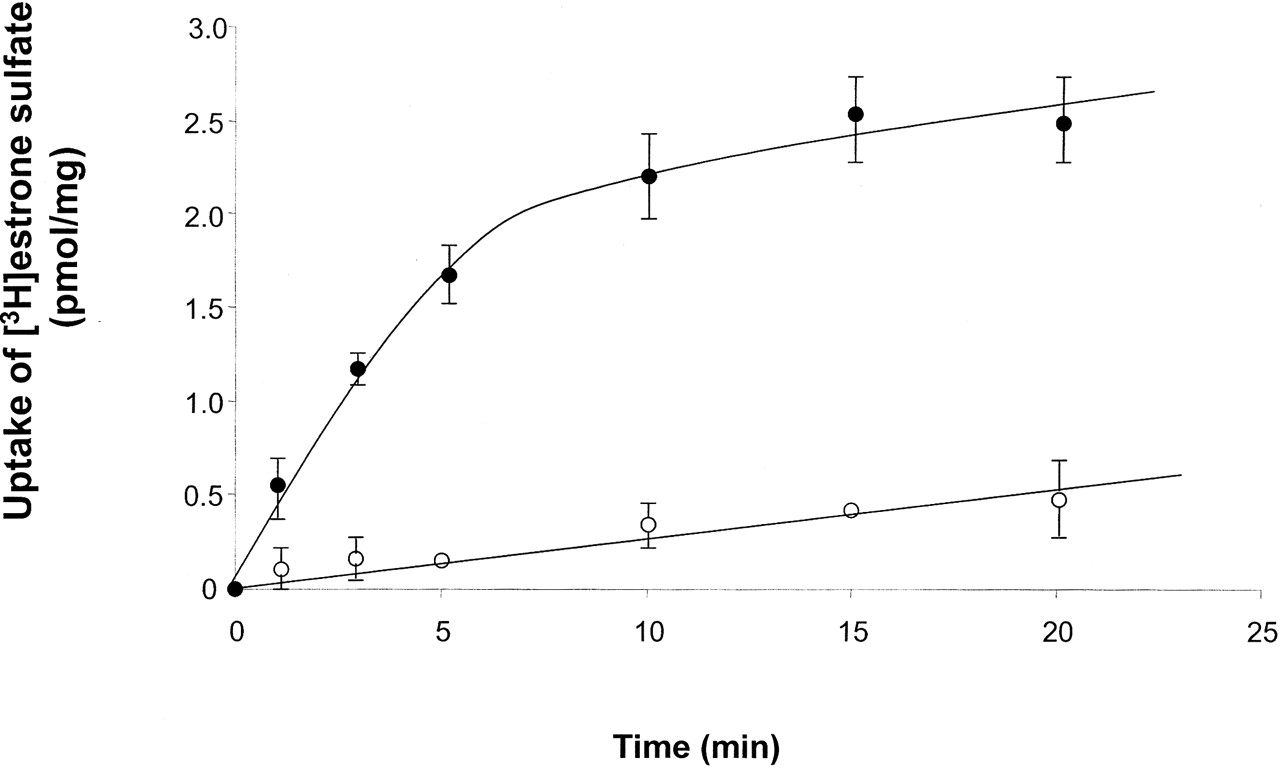
Uptake of [3H]Estrone Sulfate by Wild-Type hOAT4. Because this is the first study using COS-7 cells as an expression system for hOAT4, the functional properties of hOAT4 in these cells were characterized. The time course for [3H]estrone sulfate uptake was compared in vector-transfected (Fig. 2, ○) and hOAT4-transfected (Fig. 2, •) cells. As expected, uptake was markedly faster into hOAT4-expressing cells. In hOAT4-expressing cells, uptake increased linearly for approximately 5 min and reached a steady state between 10 and 20 min. Therefore, an uptake period of 3 min (initial rate) was chosen for future studies.

Time course of [3H]estrone sulfate uptake in COS-7 cells. [3H]estrone sulfate (100 nM) was added to the cells transfected with hOAT4 (•) and vector (○). At specified time points, cells were washed twice with ice-cold PBS, lysed with 0.2 N NaOH, neutralized with 0.2 N HCl, and measured for radioactivity. Each value represents the mean ± S.D. of three experiments.
To evaluate the substrate selectivity, we examined several radiolabeled compounds in terms of whether they are taken up into COS-7 cells via hOAT4. As shown in Fig. 3, [3H]estrone sulfate (Fig. 3a) and DHEAS (Fig. 3b) were transported into the cells. No uptake of [14C]PAH was detected (Fig. 3c). hOAT4 is known to function as a Na+-independent transporter in other systems (Cha et al., 2000). Therefore, the role of Na+ in hOAT4-mediated estrone sulfate uptake in COS-7 cells was investigated (Fig. 4). This was done by examining the effect of isosmotically replacing Na+ (137 mM) in the incubation buffer with the monovalent cation choline (137 mM choline chloride). The result showed that replacement of Na+ by choline had no effect on estrone sulfate uptake, demonstrating that the transport function is Na+-independent. The above functional characteristics of hOAT4 (substrate specificity and Na+-independence) were consistent with those obtained from other systems (Cha et al., 2000), suggesting that hOAT4-transfected COS-7 cells are a valid model system for the structure-function analyses.

Substrate specificity. The uptake of [3H]estrone sulfate, [3H]DHEAS, and [14C]PAH were measured in COS-7 cells transfected with hOAT4 (solid bars) and vector (shaded bars). Each value represents the mean ± S.D. of three experiments.

Effect of extracellular Na+ on uptake of estrone sulfate. Uptake of [3H]estrone sulfate (100 nM) after a 3-min incubation was measured in the presence or absence of Na+ (Na+ was replaced by choline). Each value represents the mean ± S.D. of three experiments.
Uptake of [3H]Estrone Sulfate by Glycine Mutants. To explore the functional role of the evolutionarily conserved glycine residues, we mutated each glycine (at positions 11, 241, 383, 388, 400, and 466) to serine. The functional properties of these mutants were then determined by measuring the uptake of [3H]estrone sulfate in mutant-transfected cells. As shown in Fig. 5, mutants G11S, G383S, G388S, and G466S exhibited significant transport activities compared with those of wild-type hOAT4. In contrast, mutants G241S and G400S almost completely lost transport function.

Uptake of [3H]estrone sulfate by cells expressing glycine mutants. Glycine (G) at positions 11, 241, 383, 388, 400, and 466 was individually mutagenized to serine (S), and the transport of 100 nM [3H]estrone sulfate in mutant-transfected cells was measured. V, pcDNA vector. Values were mean ± S.E. (n = 3). ★, P < 0.05 and ★★, P < 0.01 versus control.
To further elucidate the molecular mechanisms underlying the effects of mutations at Gly-241 and Gly-400, we mutagenized these two residues to amino acids with varying sizes of side chains, including alanine, valine, and leucine. As shown in Fig. 6, substitution of Gly-241 (Fig. 6a) and Gly-400 (Fig. 6b) with amino acids with small side chains, such as alanine, resulted in detectable transport activities, although they were much reduced compared with those of wild-type hOAT4. In contrast, substitution of Gly-241 and Gly-400 with amino acids with larger side chains, such as valine and leucine, rendered hOAT4 nonfunctional. Mutants G241S and G400S were also included in this study and shown to have little transport activity. This finding clearly demonstrates that increasingly larger side chains at positions 241 and 400 increasingly impair hOAT4 function.

The effects of glycine mutations on hOAT4 function. Glycine (G) at positions 241 and 400 was individually mutagenized to alanine (A), valine (V), and leucine (L), and the uptake of 100 nM [3H]estrone sulfate in mutant-transfected cells was measured. Mutants G241S and G400S were also included in this study. V, pcDNA vector. Values were mean ± S.E. (n = 3). ★★, P < 0.01 versus control.
Immunoblot Analysis on the Expression of Glycine Mutants. The reduced, or lack of, transport activities of Gly-241 and Gly-400 mutants could be caused by changes in the absolute number of transporters, turnover rate, substrate binding affinity, or a combination of these factors. As a first step in evaluating possible changes, we compared the protein expression levels of wild-type hOAT4 and its mutants in the total cell extracts and on the cell surface by immunoblot analysis (Fig. 7). We demonstrated above (Fig. 1) that hOAT4 in COS-7 cells is a glycosylated protein with a molecular mass of ∼83 kDa. In total cell extracts (Fig. 7, a and b, bottom), all of the hOAT4 mutants expressed a band at ∼83 kDa similar in intensity to that expressed by wild-type protein, suggesting that similar amounts of wild-type and mutant proteins are expressed in these cells. In contrast, the intensity of the 83-kDa band detected from the cell-surface pools decreased significantly in mutant-transfected cells compared with that of wild-type hOAT4-transfected cells (Fig. 7, a and b, top): the larger the side chains at positions 241 and 400, the less the expression of the mutants at the cell surface. Figure 7, c and d, shows the densitometry analyses of the surface expression for Fig. 7, a and b, respectively.

Total cell and surface expression of hOAT4 and its mutants. For total cell expression (a and b, bottom), COS-7 cells expressing hOAT4 wild type (Wt) and its mutants (G241A, G241S, G241L, G241V, G400A, G400S, G400L, and G400V) were lysed, and their proteins were separated by SDS-PAGE and visualized by immunoblot analysis using anti-hOAT4 antibody. For cell-surface expression (a and b, top), cells expressing hOAT4 wild type (Wt) and its mutants (G241A, G241S, G241L, G241V, G400A, G400S, G400L, and G400V) were biotinylated, and the labeled cell-surface proteins were precipitated with streptavidin beads, separated, and visualized as above. c and d, densitometry analyses of the intensities of the surface expression in a and b, respectively.
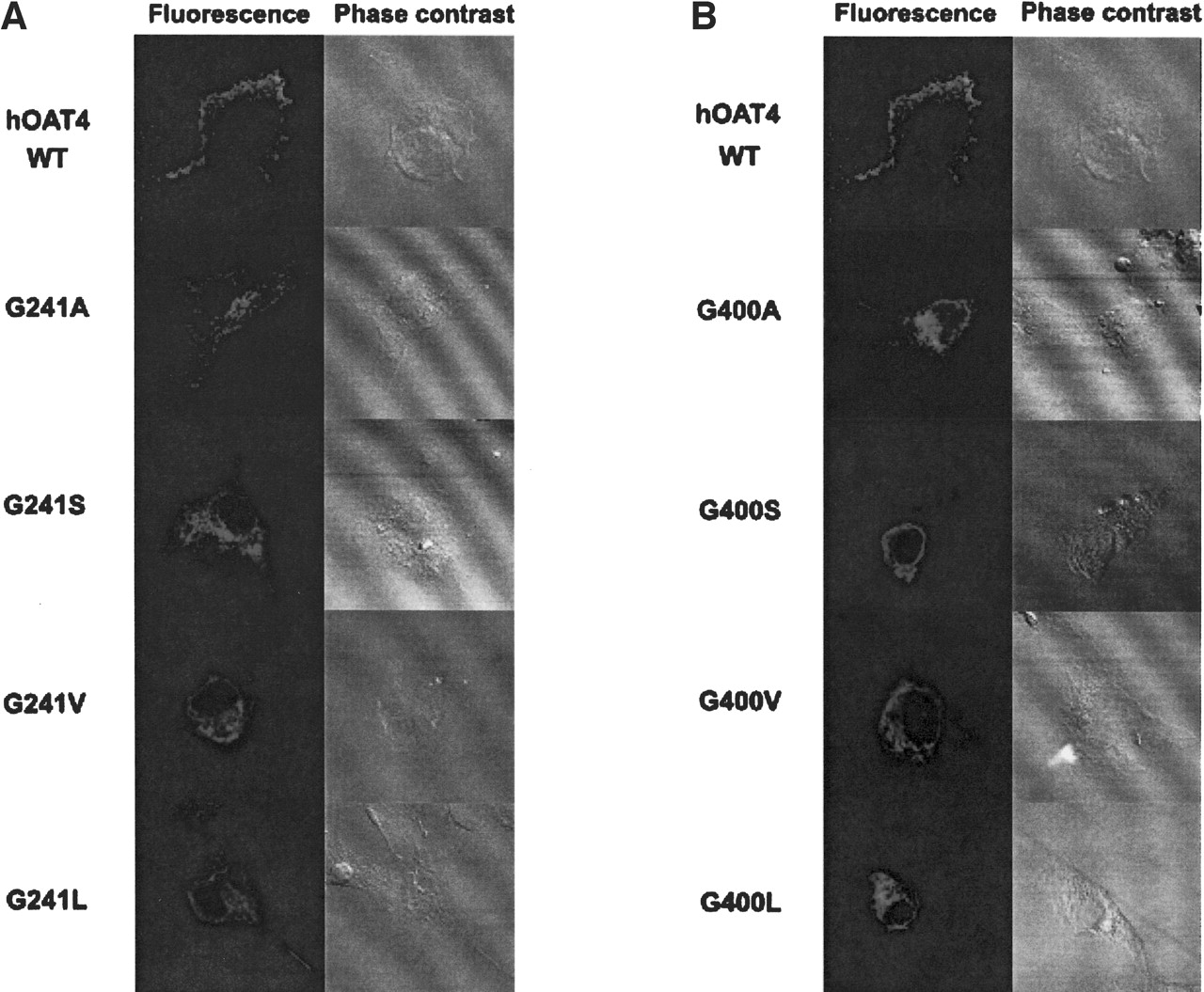
Immunofluorescence Analysis on the Expression of Glycine Mutants. Further evidence of the difficulty in transporting mutant protein to the plasma membrane was obtained by immunofluorescence (Fig. 8). Although the plasma membrane was clearly labeled (shown as green fluorescence) in cells transfected with wild-type hOAT4, fluorescence remained mainly in the intracellular compartment in cells transfected with mutants G241S, G241V, G241L (Fig. 8a) and G400S, G400V, and G400L (Fig. 8b). G241A- and G400A-transfected cells showed both surface expression and accumulation in the intracellular compartment. Phase contrast images showed that cells were fully attached to the culture dishes under all conditions. From all these data, it can be concluded that larger side chains at positions 241 and 400 impaired the proper targeting of hOAT4 to the plasma membrane.

Immunofluorescence study of the effect of substitution of Gly-241 (a) and Gly-400 (b) with larger side chains on the cellular expression of hOAT4. Wild-type hOAT4 and its mutants were expressed in COS-7 cells. The cells were then stained with anti-hOAT4 antibody and fluorescein isothiocyanate-coupled goat anti-mouse IgG. Specific immunostaining appears as bright fluorescence. Phase contrast images showed that cells were fully attached to the culture dishes under all conditions.
Kinetic Analysis of Estrone Sulfate Transport Mediated by Mutants G241A and G400A. To further examine the mechanisms underlying the reduced transport activity of hOAT4 by substitution of Gly-241 and Gly-400 with alanine, we determined [3H]estrone sulfate uptake at different substrate concentrations. An Eadie-Hofstee analysis of the derived data (Fig. 9) showed that alanine substitution of these residues resulted in decreased Vmax (53.8 ± 1.1 pmol/μg · 2 min with wild-type hOAT4, 21.2 ± 0.5 pmol/μg · 2 min with G241A, and 25.3 ± 1.7 pmol/μg · 2 min with G400A) and increased Km (3.2 ± 0.1 μM with wild-type hOAT4, 5.2 ± 0.4 μM with G241A, and 6.3 ± 0.7 μM with G400A).

Kinetic analysis of estrone sulfate transport mediated by mutant G400A. Kinetic characteristics were determined at substrate concentrations ranging from 0.1 to 10 μM (2-min uptake) using cells expressing hOAT4 wild type (Wt; closed circles; a and b), mutant G241A (open circles; a), and mutant G400A (open circles; b). Transport kinetic values were calculated using the Eadie-Hofstee transformation. Values were mean ± S.E. (n = 3).
Discussion
The vital importance of organic ion transport in body drug disposition is clear (Dresser et al., 2001; You, 2002; Koepsell et al., 2003). However, our knowledge about the structure-function relationships of organic ion transporters is minimal. The aim of this study was to identify the structural determinants important for the function of hOAT4. To this end, we explored the roles of six glycine residues (at positions 11, 241, 383, 388, 400, and 466) in hOAT4. These glycine residues are conserved across organic ion transporters of different origin, from human to C. elegans. This led to our hypothesis that they may play important roles in the function of these transporters.
By first mutagenizing each of these glycine residues to serine, we found that mutants G11S, G383S, G388S, and G466S transported significant amounts of estrone sulfate. In contrast, mutants G241S and G400S lost ability to transport this molecule, suggesting an important role of Gly-241 and Gly-400 in hOAT4 function. To further investigate the significance of Gly-241 and Gly-400, we then mutagenized these two residues to several other amino acids with varying side chains, such as alanine, valine, and leucine. We showed that the size of the side chain of the residues at positions 241 and 400 is of functional significance provided no charge is present. When native Gly-241 and Gly-400 were replaced with alanine, a residue with a side chain slightly larger than that of glycine and equally neutral, the resulting hOAT4 molecules (G241A and G400A) still exhibited measurable transport activities, although much reduced compared with that of wild-type hOAT4. In contrast, when native Gly-241 and Gly-400 were replaced with valine or leucine, residues with consecutively larger side chains than alanine and devoid of charge, the resulting hOAT4 molecules (G241V, G241L, G400V, and G400L) became nonfunctional.
The reduced or lack of transport activity of glycine mutants could result from the reduced or lack of expression of the mutant tansporter protein or could result from impaired binding abilities of the mutants for their substrates. By directly measuring both total cell and cell surface expression of these mutants, we showed that despite the similar total cell expression of these mutants relative to that of wild-type hOAT4, larger side chains at positions 241 and 400 resulted in a decrease in the expression of the mutants at the cell surface. These results suggest that substitution of Gly-241 and Gly-400 with larger side chains does not interfere with the total cell expression of the transporter protein but rather interferes with the trafficking of the transporter to the plasma membrane. Our immunofluorescence study confirmed these results. In addition to the much lower cell-surface expression, our kinetic analysis showed that the reduced transport activity was also contributed to by a reduced binding affinity of the mutants for its substrates. Substitution of Gly-241 and Gly-400 with alanine resulted in a 1.6- to 2-fold increase in Km compared with that of wild-type hOAT4. Based on the predicted 12-transmembrane topology of hOAT4, Gly-241 and Gly-400 reside in transmembrane domain 5 and at the mouth of transmembrane domain 8, respectively. Glycine has been implicated to play an important role in stabilizing transmembrane helix-helix association for membrane proteins such as water channel aquaporin-1 (Murata et al., 2000; Russ and Engelman, 2000). Therefore, our data suggest that transmembrane domains 5 and 8 in hOAT4 may form part of the translocation pathway for the substrates.
In conclusion, mutation of glycine at positions 241 and 400 impairs hOAT4 function by impairing both the trafficking of the transporter to the cell surface and its binding ability for the substrates. The extent of the impairment is dependent on the size of the side chains at positions 241 and 400. The increasingly larger side chains increasingly impair hOAT4 function. This is the first identification and characterization of critical amino acid residues in hOAT4 and may provide important insights into the structure-function relationships of the organic ion transporter family.
Footnotes
-
This work was supported by National Institutes of Health Grant R01-DK 60034 (to G.Y.).
-
ABBREVIATIONS: OAT, organic anion transporter; OCT, organic cation transporter; hOAT4, human organic anion transporter 4; DHEAS, [3H]dehydroepiandrosterone 3-sulfate; [14C]PAH, [14C]para-aminohippurate; PCR, polymerase chain reaction; PBS, phosphate-buffered saline; NHS-SS-biotin, sulfosuccinimidyl 2-(biotinamido)-ethyl-1,3-dithiopropionate; PAGE, polyacrylamide gel electrophoresis; PVDF, polyvinylidene difluoride.
- Received October 8, 2003.
- Accepted February 2, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}