Article Text

Statistics from Altmetric.com
Genomic imprinting is a phenomenon whereby genes are differentially expressed according to their parental origin.1,2 Mutations which affect the epigenetic states of imprinted domains underlie a number of diseases, including developmental abnormalities, malignant tumours, and psychiatric diseases.3 To date, >50 imprinted genes have been identified in human and mouse (www.otago.ac.nz/IGC). For some genes, imprinting has been shown to be tissue specific4–9 and/or developmentally regulated.10,11 For others, polymorphic imprinting (the term for the imprinting of a gene that is variable between individuals) has also been observed.12,13 These variations in gene expression are considered to be a source of phenotypic heterogeneity in human disease with parent-of-origin effects.4
Three imprinted loci have been identified on human chromosome 7.14 One of the loci is located on 7q21.3 and two genes, SGCE and PEG10, are known to be imprinted (fig 1A). Initially the mouse Sgce gene (encoding sarcoglycan-epsilon) was identified as a maternally imprinted (paternally expressed) gene in a subtractive screening experiment using cDNA libraries derived from parthenogenetic and wild-type fibroblast lines.15 The human SGCE gene has also been confirmed to be imprinted.16–18PEG10 (paternally expressed gene 10), which is immediately telomeric to SGCE, was also shown to be imprinted in embryonic villi.19 Moreover, the mouse Asb4 gene (ankyrin repeat and SOCS box-containing protein 4), which is 800 kb telomeric to Sgce, was found to be maternally expressed by cDNA micro-array based high-throughput screening for genes differentially expressed between mouse parthenogenetic and androgenetic embryos.6 The imprinting status of the human ASB4 and other genes adjacent to SGCE/PEG10 has not yet been determined, but systematic imprinting analyses of mouse genes indicate that the 1 Mb interval encompassing Sgce/Peg10 is part of a large imprinted locus; Peg10 was confirmed to be paternally expressed, and neurabin (Ppp1r9a), Pon2, and Pon3 were found to be preferentially maternally expressed.20 The mouse calcitonin receptor gene (Calcr), which is located 1.2 Mb centromeric to the Sgce/Peg10 locus, was recently shown to be imprinted in a brain specific manner.9 Moreover, DLX5, a gene located 2.4 Mb telomeric to the SGCE/PEG10 locus, is imprinted in human lymphoblasts and brain tissue.21 These findings raise the possibility that the cluster of imprinted genes identified in mouse (Sgce to Asb4) could extend further in both centromeric and telomeric directions.

(A) A 1.2 Mb interval containing three imprinted genes and PPP1R9A on human chromosome 7q21.3. Genes shown to be imprinted in human are highlighted in green (paternally expressed) or red (maternally expressed). Red asterisks indicate the genes whose mouse orthologue is shown to be preferentially maternally expressed, but whose patterns in human have yet to be determined. (B) Exon–intron structure of the human PPP1R9A gene and SNPs (single nucleotide polymorphisms) used for allelic expression analysis. Two alternative spliced isoforms I and II are shown. Only the size of exons are drawn to scale (the gene encompasses 385 kb as indicated in panel A). SNPs used for allelic expression experiments are marked by asterisks. The positions of DNA fragments amplified by RT-PCR and sequenced for allelic expression patterns are shown by black bars.
The human PPP1R9A gene encoding neurabin I is adjacent to SGCE, PEG10, and ASB4 (fig 1A). Neurabin I (neural tissue specific F-actin-binding protein I) was originally purified from rat brain and has been characterised as a key candidate molecule in synaptic formation and function.22 It has also been shown that neurabin I is highly concentrated in the synapse of developed neurons and is involved in neurite formation. The deduced 1098 amino acid sequence for rat neurabin I contains an F-actin-binding domain, a PDZ domain, and coiled-coil domains.22 As the name indicates, the expression of the gene and protein was shown to be brain specific in adult rat tissues.22,23 The gene symbol is based on “protein phosphatase 1, regulatory (inhibitor) subunit 9A” because neurabin I has been shown to bind protein phosphatase I (PP1) and inhibit its activity.24 The PP1/neurabin I complex has been demonstrated to regulate actin cytoskeleton dynamics.25 Furthermore, molecules implicated in the control of neuronal morphology, such as p70S6K23 and kalirin-7 (a GTP-exchange factor),26 have been identified to bind the PDZ domain of neurabin I. Lastly, the C-terminus of neurabin I binds the trans-Golgi protein TGN38, suggesting it is a multi-functional molecule that regulates membrane and cytoskeletal functions.27
Key points
-
Deregulation of imprinted genes has been observed in various human diseases. The human PPP1R9A gene, which encodes neurabin I, is located in a cluster of imprinted genes on human chromosome 7q21. The protein has been shown to be a regulatory subunit of protein phosphatase I, and controls actin cytoskeleton reorganisation.
-
The expression patterns of human and mouse PPP1R9A gene were analysed by Northern blot and RNA in situ hybridisation. Allelic expression patterns were determined by DNA sequencing and a single nucleotide primer extension method (SNaPshot assay). PPP1R9A was found to be transcribed in both neuronal and multiple embryonic tissues. In allelic expression analyses in human and mouse, the gene was found to be preferentially transcribed from the maternally inherited chromosome mainly in embryonic skeletal muscle tissues, but was biallelically expressed in other embryonic tissues. Preferential or exclusive maternal expression was also observed in extra-embryonic tissues.
-
Our findings suggest that PPP1R9A is not neuronal specific as shown previously, but rather important for early development of multiple tissues, some being influenced by the imprinting status. Dosage control of neurabin I through imprinting mechanisms may be necessary for proper skeletal muscle differentiation, which requires reorganisation of the actin cytoskeleton.
The mouse Ppp1r9a was shown recently to be preferentially maternally expressed in extra-embryonic tissue, but biallelically in neonatal tissue.20 Independently, we were conducting imprinting analyses for PPP19A since imprinted genes are often clustered, making PPP1R9A an obvious positional candidate to study. Moreover, since some imprinted genes are thought to be involved in mental dysfunction28 and maternal nurturing behaviour,29 we aimed to determine if PPP1R9A could be related to neurobehavioural disorders with parent-of-origin effects associated with chromosome 7. Intriguingly, PPP1R9A was found to be imprinted (maternally expressed) mainly in skeletal muscle tissues, but not in brain. Ppp1r9a was also imprinted in extra-embryonic tissues in mouse. Taken together, our data indicate that neurabin I is not neuronal specific as previously described and its differential expression could play an important role in multiple tissues.
METHODS
mRNA sequence for mouse Ppp1r9a
The exon–intron structure of the mouse Ppp1r9a gene was estimated by the alignment of human mRNA sequences (AK096514 and AB033048) with mouse genomic DNA by BLAT (http://genome.ucsc.edu). PCR primers mNeu-F (5′-ATGAGCATTGGCTTTTCACC-3′) on exon 2 and mNeu-R 5′-GGCGGGACTTCTCTCAAACT-3′ on exon 16 were used in RT-PCR to amplify cDNA fragments containing the entire open reading frame for the mouse Ppp1r9a gene. Random-primed cDNA from total RNA of 15.5 day post coitum (dpc) brain was used as template DNA with PfuUltra High-Fidelity DNA Polymerase (Stratagene). The cycling conditions were: initial denaturation at 94°C for 3 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 4 min. The amplified products (3.4 kb in size) were subcloned into the pCR-Script vector (Stratagene) and sequenced.
Northern blot analysis
A human MTN blot (Clontech, 7760-1) containing poly A RNA (2 μg per lane) from human adult tissues was hybridised with 32P[α]dCTP-labeled probe (187 bp fragment corresponding to nt 3063–3249 of GenBank acc. number AB033048). The hybridisation and washing conditions were described previously.30
RNA in situ hybridisation
A 447 bp fragment corresponding to nt 3857–4303 of GenBank acc. number AY308065 was subcloned into pCR-Script vector (Stratagene) and used as template DNA for sense/antisense riboprobes. Probes were hybridised with paraffin-embedded sagittal sections of 15.5 dpc embryo (from the CD1 strain). The conditions used for hybridisation, washing, and signal detection were described previously.31
Allelic expression analysis on human fetal samples
A 201 bp DNA fragment was amplified from fetal genomic DNA using primers ex161F/1R (5′-CACTGGCAAACACAACACAG-3′ and 5′-GTCCTTTGGAAAGTGGATAACC-3′), and was sequenced to determine the genotype for SNP rs854541 on exon 16. Primers AB033048-1813F/3249R (5′-GCCTGGATATGATAGATGACGA-3′ and 5′-CCACCTGGGAAATCTTGAGT-3′) were used to amplify a 1437 bp fragment (for isoform I) from fetal cDNAs. The cycling conditions were: initial denaturation at 94°C for 3 min, followed by 38–40 cycles of denaturation at 94°C for 30 s, annealing at 58°C for 30 s, and extension at 72°C for 90 s. AB033048–3249R was used as a sequencing primer.
A 499 bp DNA fragment was amplified from fetal genomic DNA using primers 12b-1F/1R (5′-CTCAGTGGCATAACCTTCCTT-3′ and 5′-ACATGGAGGGGTTGTTTCTC-3′) and sequenced to determine the genotype for SNP rs854524 on exon 12b. Primers AK000075-174F/632R (5′-CCCTTCTCGTCTGACCACAT-3′ and 5′-TTTCCGGCTGTCATTAAACC-3′) were used to amplify cDNA products (459 bp) for isoform II. The cycling conditions were: initial denaturation at 94°C for 3 min, followed by 35–38 cycles of denaturation at 94°C for 30 s, annealing at 58°C for 30 s, and extension at 72°C for 45 s. AK000075–524R was used as a sequencing primer.
The PCR products were purified using microCLEAN (Microsone) and were sequenced directly using Big Dye terminator kit and resolved on a 310 Genetic Analyzer (Applied Biosystems).
To determine the allelic expression pattern of the PPP1R9A by DNA sequencing, the region containing the peaks for the SNP sites was enlarged on imaging software, and the heights of the two alleles (A and B) were measured. When the peak ratio (A/B) in a cDNA sample was between 50 and 200% compared with that of heterozygous genomic DNA control, PPP1R9A was regarded as being biallelically expressed.
Allelic expression analysis on mouse embryonic samples
Total RNA was extracted from embryonic tissues (12.5 dpc, 15.5 dpc, and postnatal (P0)) from reciprocal F1 crosses between C57BL/6J and CAST/Ei. cDNA was synthesised using random primers. Primers 441F (5′-TCACGGAGACTCGAAAGATGT-3′) and 1471R (5′-GTCGTCATTTCTCCGGTCAT-3′) were used for RT-PCR to amplify a 1031 bp fragment containing SNPs in exon 2 (nt 536 and 616 of GenBank acc. number AY308065), followed by direct sequencing by primer 759R (5′-TTGGTGACGGTAACAGATGG-3′).
Quantitative allelic expression analysis (SNaPshot assay)
PCR products were purified by QIAquick Gel Extraction Kit (Qiagen). Purified PCR product (0.01–0.40 pmol) was used for SNaPshot primer extension reaction. A 0.2 μM of primer and 2 μl SNaPshot Ready Reaction Mix containing fluorescence-labelled ddNTPs, Amplitaq DNA polymerase, and reaction buffer (Applied Biosystems) were added to a total volume of 10 μl. The extension primers used were 5′-TGTTCAATTTCTTGGGTTAGGT-3′ for rs854541 (PPP1R9A isoform I), 5′-AAACCACCAACAAGAAAATATTAC-3′ for rs854524 (PPP1R9A isoform II), and 5′-GAGCCCCCGGACGGAAGC-3′ for nt 616 of AY308065 (mouse Ppp1r9a). The cycling conditions for the reaction were 25 cycles at 95°C for 10 s, 58°C for 5 s, and 60°C for 30 s. One unit of shrimp alkali phosphatase was added, and the mixture was incubated at 37°C for 1 h, followed by 75°C for 15 min. The reactions were resolved on an ABI 3100 Genetic Analyzer, and signal intensities for two alleles at an SNP site were measured using GeneScan Analysis Software Version 3.5. The percentage ratio of the two alleles at SNP sites for cDNA samples was corrected using the mean of the ratios obtained from analyses of three heterozygous genomic DNA samples as described previously.32
DNA methylation analysis
Genomic DNA was isolated from normal, mUPD7, and pUPD7 lymphoblastoid cells. The mUPD7 and pUPD7 patients had undergone microsatellite repeat analysis to confirm uniparental disomy for the entire length of chromosome 7.33 Tissue specific methylation was analysed in fetal liver, fetal muscle, and placenta. A 15 μg sample of genomic DNA was pre-digested with XbaI, then with the methylation-sensitive HpaII or Hin6I, or the HpaII methylation-insensitive isoschizomer MspI, before electrophoresis and Southern blot transfer to Hybond N+ membrane. The filters were probed with a 1.3 kb PCR product (nt 59398–60715 of AC073886). Filters were pre-hybridised (0.2 M NaPO4 pH 7.2, 1 mM EDTA, 1% BSA, 7% SDS, 15% deionised formamide) for a minimum of 3 h/65°C, and hybridised with a radio-labelled probe overnight. Filters were washed three times (40 mM NaPO4, 1 mM EDTA, 1% SDS) for 7 min/65°C. Blots were exposed to x ray film with intensifying screens at −70°C for 1 week.
RESULTS
Gene structure and expression pattern of the human and mouse PPP1R9A gene
GenBank acc. number U72994 represents the mRNA sequence containing the entire open reading frame of the rat neurabin I protein (1095 amino acids).22 The human mRNAs, AK096514 and AB033048, were found to contain the entire open reading frame for the neurabin I protein (1098 amino acids), which showed 90% sequence homology with the rat protein sequence. Alignment of the human mRNA sequences with genomic DNA sequence revealed that human PPP1R9A is composed of 16 exons encompassing 385 kb of DNA. We designated this exon–intron structure to be isoform I (fig 1B). A partial mRNA sequence (AK000075) was found to contain additional exons (12b–12e) between exon 12a and 13, but it lacked exon 14 (fig 1B). We have, therefore, designated this as isoform II and could confirm by RT-PCR that it shared the exon–intron structure with isoform I for the rest of exons (data not shown). ESTs AW205092 and BG183318 were found to contain an alternative form of exon 12e (named 12e′), which is 115 bp longer because of an alternative splicing donor site (this was designated PPP1R9A isoform III). We determined the mRNA sequence for the mouse Ppp1r9a gene (GenBank acc. number AY308065) by sequencing RT-PCR products containing the entire open reading frame (exons 2–16, see Methods).
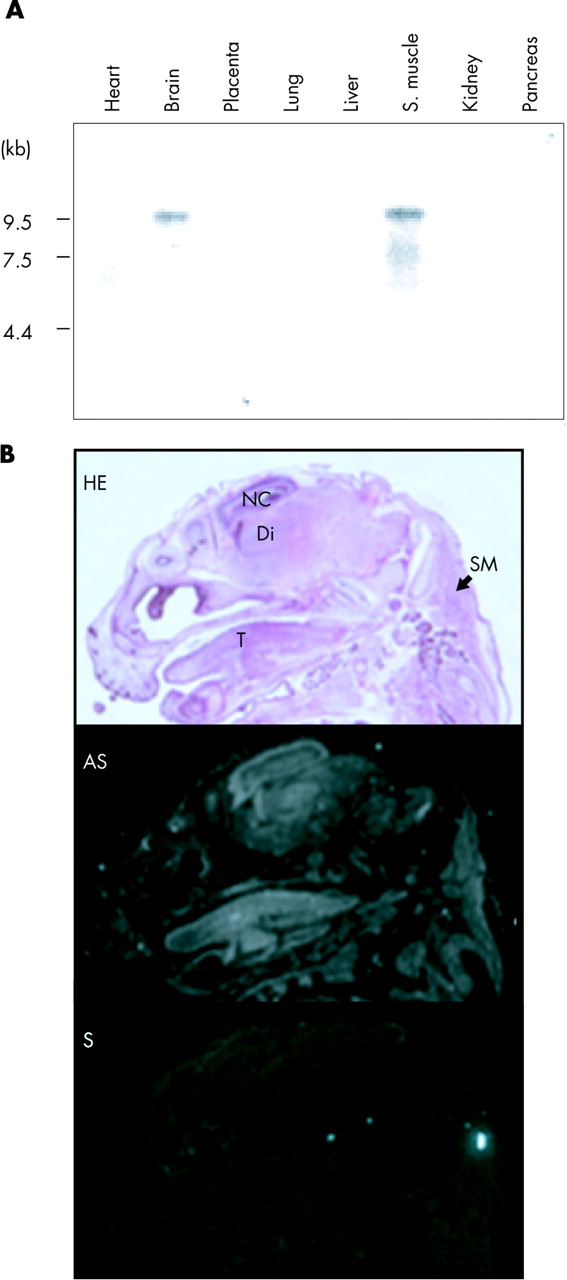
Previously, neurabin I was shown to be expressed only in the brain among rat adult tissues.22,23 However, our Northern blot analysis detected a 9.5 kb band in the brain and skeletal muscle among eight human adult tissues examined (fig 2A). The mouse gene has been shown to be expressed in adult skeletal muscle.34 Therefore, the expression of neurabin I in skeletal muscle may be age-dependent in the adult stage of these species. Alternatively, developmental regulation for the gene in skeletal muscle tissues may be different among species.
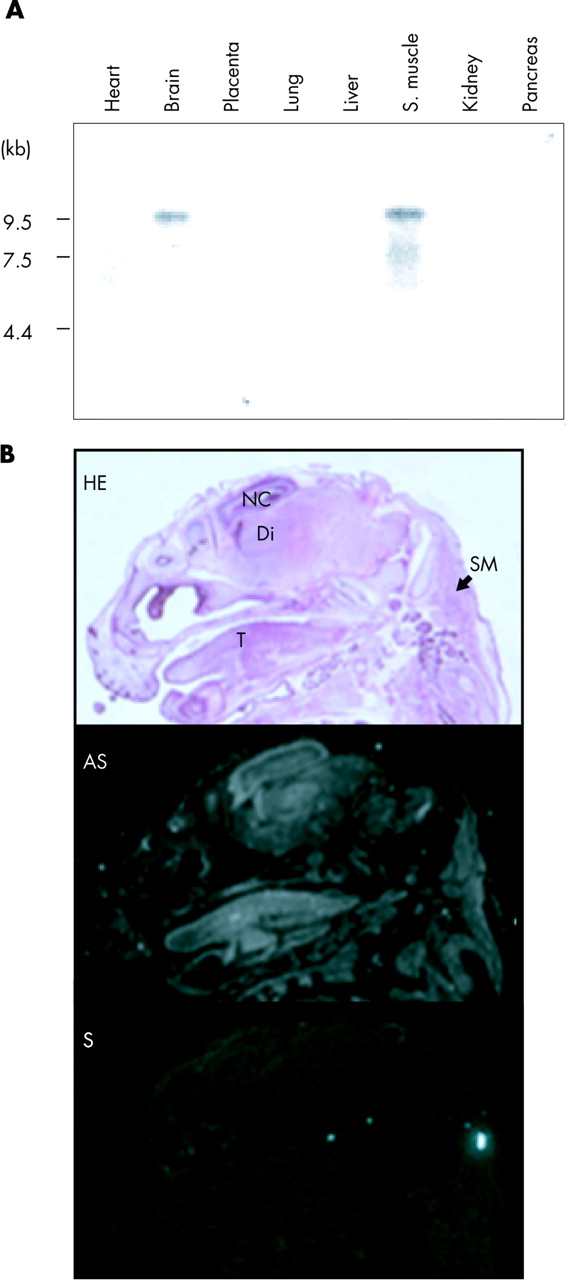
(A) Northern blot analysis of the human PPP1R9A gene on human adult tissues. Northern blot data demonstrate expression in brain and skeletal muscle. (B) RNA in situ hybridisation for the mouse Ppp1r9a gene on the head of 15.5 dpc mouse embryo. The top panel shows a haematoxylin-eosin (HE) stained section. Di, diencephalons; NC, neopallial cortex; SM, skeletal muscle at the cervical region; T, tongue. The middle and bottom panels show the patterns of hybridisation signals detected by antisense (AS) and control sense (S) probes, respectively.
The expression of Ppp1r9a in embryonic development was also examined by RNA in situ hybridisation. At 15.5 days post coitum (dpc), Ppp1r9a is highly expressed in the brain and the skeletal muscle (fig 2B). In the brain, Ppp1r9a is strongly expressed in the neopallial cortex (future cerebral cortex) and diencephalon (thalamus). The gene is also expressed in the skeletal muscle, in the developing tongue, in the cervical region, and around the primordium of the costal and scapula cartilages (data not shown).
Allelic expression analysis in human fetal tissues
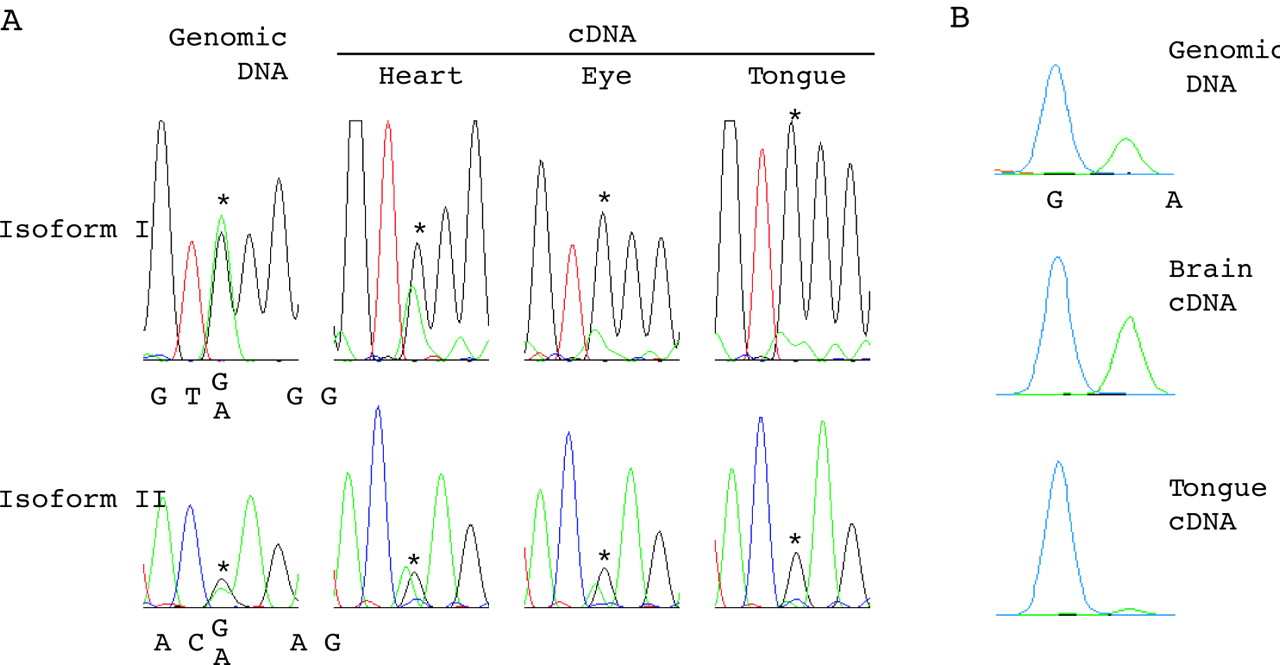
Alternatively spliced isoforms I and II of human PPP1R9A were examined for their imprinting status. Single nucleotide polymorphisms (SNPs) rs854541 in exon 16 and rs854524 in exon 12b were used for allelic expression analysis for isoforms I and II, respectively (fig 1B). Isoform III was excluded from the analysis because of its low expression level (data not shown). Eight and six fetuses were found to be heterozygous for the SNPs in isoforms I and II, respectively (table 1). The allelic expression pattern of PPP1R9A isoforms in 10 types of tissues from those fetuses was initially assessed by direct sequencing of RT-PCR products, and preferential or monoallelic expression was observed in some tissues (fig 3A). The observed biased allelic expression was confirmed to be reproducible in at least two independent experiments. To quantitate the allelic expression ratio, the same fetal cDNA samples (if available) were subjected to SNaPshot assay (Applied Biosystems), which is a fluorescence-based primer extension method and has been applied for quantitative allele frequency measurement for SNPs.32,35,36 Examples of SNaPshot outputs are shown in fig 3B. Table 1 summarises the percentage ratio of the two alleles determined by SNaPshot assay or allelic expression pattern determined by DNA sequencing. Monoallelic expression in the tongue of fetus IC-77 for both isoforms, and preferential expression in the eye of fetus IC-77 for isoform I were detected by DNA sequencing (fig 3A). The allelic expression pattern for the remaining tissues from IC-77, the tissues from IC-46 and IC-58, and the brain from IC-56 were determined to be biallelic (see Methods for the criterion). Although determination of the allelic expression pattern by DNA sequencing is not quantitative, our results clearly showed that PPP1R9A is monoallelically expressed in tongue.
Allelic expression analysis for the human PPP1R9A isoforms

Allelic expression analysis for the human PPP1R9A isoforms. (A) Electropherograms for the sequencing of genomic DNA and cDNAs (heart, eye, and tongue) for fetus IC-77 are shown. Upper and lower panels are for isoforms I and II, respectively. Asterisks show the positions of SNPs (rs854541 for isoform I and rs854524 for isoform II). IC-77 was heterozygous for both SNPs. Biallelic or biased allelic expression was observed in the tissues examined. (B) Examples of quantitative SNaPshot data. The three panels are for SNP rs854541 (isoform I) on genomic DNA, brain cDNA, and tongue cDNA for fetus IC-62. The blue and green peaks represent G and A alleles, respectively. Note the unequal heights for the two alleles on genomic DNA. Unequal allelic detection for heterozygous SNP sites has been frequently observed.36–38 This is considered to be due to background and other factors influencing peak heights (such as differential efficiencies of the incorporation of the ddNTPs and the unequal emission energies of the different fluorescent dyes). The allelic expression ratios for cDNA samples were corrected by the equation used elsewhere.36
Mono-allelic or preferential expression was detected mainly in skeletal muscle tissues (limb and tongue) and also in eye and placenta (but with a lower frequency) (table 1). The maternal allele was exclusively or preferentially expressed in three informative cases (fetus IC-37, 56, and 77).
DNA methylation analysis in human lymphoblastoid cell lines and fetal tissues
A CpG island overlapping with exon 1 of PPP1R9A was found using the default setting (GC >50%, length >200, Obs CpG/Exp CpG >0.6) of the CpG island prediction algorithm in the UCSC Genome Browser (http://genome.cse.ucsc.edu). The CpG island is 627 bp in length (83 CpG dinucleotides; %CG = 73.6) and corresponds to nt 59785–60411 of GenBank acc. number AC073886. Ten HpaII and 12 Hin6I methylation-sensitive restriction sites were found in the CpG island. The methylation status of these sites was examined in lymphoblastoid cell lines derived from individuals with bi-parental inheritance or uniparental disomy of chromosome 7 (UPD7) and fetal tissues (placenta, liver, and muscle) by Southern blot hybridisation. A 1.3 kb probe encompassing the entire CpG island was hybridised with blots containing genomic DNA digested with XbaI only, XbaI and Hin6I, XbaI and HpaII, and XbaI and MspI (the methylation-insensitive isoschizomer of HpaII) (fig 4). The same band pattern was detected on genomic DNA derived from paternal and maternal UPD7 lymphoblastoid cell lines, suggesting that the CpG island is not differentially methylated between two parental alleles in lymphoblastoid cell lines. No evidence for differential or tissue specific methylation was observed in the three fetal tissues tested, including muscle, where imprinted expression was detected.

DNA methylation analysis for the CpG island located at the 5′ end of the human PPP1R9A gene by Southern blot hybridisation. A 1.3 kb probe encompassing a CpG island overlapping with exon 1 of PPP1R9A was hybridised with blots containing genomic DNA digested with XbaI only, XbaI and Hin6I, XbaI and HpaII, and XbaI and MspI. Genomic DNA samples used were prepared from three lymphoblastoid cell lines (normal, paternal UPD7, and maternal UPD7) and three fetal tissues (placenta, liver, and muscle). The approximate band sizes are shown on the right.
Imprinting analysis in mouse embryonic tissues from reciprocal F1 hybrids
The imprinting pattern of the mouse orthologue (Ppp1r9a) was examined by DNA sequencing and SNaPshot assays in embryonic and extra-embryonic tissues of interspecific reciprocal crosses between C57BL/6J (B6) and CAST/Ei. F1 hybrids between female B6 and male CAST, and female CAST and male B6 are designated as BxC and CxB, respectively. SNPs between B6 and CAST were identified in exon 2 (nt 536 and 616 of acc. number AY308065) and exon 16 (nt 3980, 4124, and 4381 of acc. number AB091828). Consistent with the exon–intron structure of the human gene, RT-PCR using primers spanning exon 12a to exon 16 for the mouse Ppp1r9a gene detected spliced isoforms in some of the tissues tested (data not shown). The allelic expression pattern of the mouse gene was primarily assessed by sequencing a 1031 bp cDNA fragment (nt 441–1471 of GenBank acc. number AY308065) that contains the two SNPs in exon 2 and spans exons 2–4. The RT-PCR products for the region are presumably amplified from multiple alternatively spliced isoforms between exon 12a and exon 16. However, since no isoform specificity was observed in the imprinting pattern of the human gene, allelic expression analysis for each spliced isoform was not performed for the mouse gene.
Consistent with the allelic expression pattern observed for the human PPP1R9A, preferential maternal expression was detected only in limb and tongue, most clearly in the postnatal stage (P0), among the nine embryonic tissues examined (table 2). The allelic expression pattern of Ppp1r9a in heart (12.5 and 15.5 dpc), intestine (15.5 dpc), and skin (15.5 dpc) from BxC and CxB hybrids was assessed by DNA sequencing only and found to be biallelic (data not shown). Brain and tongue from reciprocal F1 hybrid embryos between B6 and JF1/Ms (designated as BxJ and JxB) were also analysed using an SNP at nt 616 of acc. number AY308065, and preferential maternal expression of Ppp1r9a was observed in tongue, but not in brain. Yolk sac was found to show the most striking maternal specific expression in the mouse tissues analysed; exclusive maternal expression was detected at 15.5 dpc in BxC hybrids (fig 5 and table 1). Placenta also demonstrated preferential maternal expression, although we cannot exclude the possibility that the observed maternal preferential expression is due to contamination of maternal decidua.
Allelic expression ratio of the maternal (M) and paternal (P) alleles of the mouse Ppp1r9a gene in F1 hybrid tissues

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Allelic expression analysis for the mouse Ppp1r9a gene. (A) Electropherograms for sequencing of genomic DNA for an SNP in exon 2 of Ppp1r9 are shown. Nt 616 of GenBank acc. number AY308065 for mouse Ppp1r9a mRNA was found to be polymorphic between B6 and CAST strains. B6 and CAST were homozygous for C allele and A allele, respectively. As a result, BxC and CxB hybrids were heterozygous for the SNP site (only the result from BxC hybrid is shown). (B) Electropherograms for sequencing of RT-PCR products (for brain, tongue, and yolk sac) are shown. Embryonic tissues from three developmental stages (12.5 dpc, 15.5 dpc, and postnatal (P0)) were analysed. The expression ratio of maternal allele (C allele in BxC and A allele in CxB) determined by SNaPshot assay is shown for each sample.
The degree of maternal specific expression was developmental stage specific in skeletal muscle tissues and yolk sac, being markedly greater during the latter developmental stage. Imprinting was found to be more relaxed when the paternal allele was derived from B6 than from CAST or JF1. This tendency was consistent for all developmental stages in all four tissues in which preferential maternal expression was detected (fig 5 and table 2). For all tissues where monoallelic or preferential expression was observed, at least two independent embryos were subjected to allelic expression analyses. The extent of mono-allelic/preferential expression was reproducibly similar between the same type/stage of tissues from different embryos.
DISCUSSION
Our finding that Ppp1r9a is imprinted in placenta and yolk sac fits well with the idea that parent-of-origin specific gene expression is most often observed in extra-embryonic tissues8 and is consistent with the results of Ono et al.20 However, the pattern of tissue specific imprinting observed in embryonic tissues is intriguing. This is the first report of skeletal muscle specific imprinting in embryonic tissues conserved between human and mouse. Biased allelic expression was also observed in some of the human fetal eye samples, but not in the eye samples from 15.5 dpc mouse embryos. Ppp1r9a may be imprinted in a limited type of ocular cells at a certain developmental stage; allelic expression analyses specific to each cell type from eye would be required to assess such a possibility. It is also possible that Ppp1r9a is imprinted in other tissues or other developmental stages that were not analysed in this study.
The extent of preferential maternal expression of Ppp1r9a was found to be strain-dependent in mouse. Imprinting was more relaxed when the paternal allele was derived from C57BL/6J (B) than from CAST/Ei (C) or JF1/Ms (J) in all tissues and stages where preferential maternal expression was detected. Variations in the extent of preferential expression were also observed in humans (table 1). Therefore, in both humans and mice, differences in genetic background seem to affect the imprinting regulation of PPP1R9A (polymorphic imprinting). Strain-dependent relaxation of imprinting has been reported in the mouse Kvlqt1 gene in reciprocal crosses between 129/Sv and CAST/Ei.11
Although the number of genes identified to demonstrate tissue specific imprinting is increasing,4–9 the molecular mechanisms underlying this phenomenon are largely uncharacterised. Our DNA methylation analysis for the CpG island overlapping with exon 1 of PPP1R9A in human lymphoblastoid cell lines and fetal tissues did not reveal any evidence of differential methylation in a parent-of-origin or a tissue specific manner. The methylation status of the CpG island at the 5′ end of the mouse Ppp1r9a was examined in mouse 10 dpc embryo and placenta, and the CpG island was also not found to be methylated.20 These results indicate that the tissue specific maternal expression of the human and mouse PPP1R9A gene is not regulated by the methylation status of the CpG island at exon 1.
Yamasaki et al7 demonstrated cell type specific imprinting of the Ube3a gene. Ube3a is expressed maternally in neurons, but biallelically in glial cells, whereas the antisense transcript overlapping with Ube3a was expressed only in neurons and only from the paternal allele. Reciprocal imprinting of sense and antisense transcripts present only in neurons suggests that the neuron specific imprinting mechanism is related to the lineage determination of neural stem cells.7 Similar to the developmental regulation of imprinting of Ube3a in the neuronal lineage, the preferential maternal expression of Ppp1r9a in skeletal muscle tissue was found to become more prevalent at later developmental stages. This suggests that the imprinting mechanism for Ppp1r9a in skeletal muscle tissues may be related to the commitment of myoblast differentiation.
In skeletal muscle differentiation, the fusion of mono-nucleated myoblasts to form multi-nucleated myotubes is a prominent event requiring reorganisation of actin cytoskeleton.37 Furthermore, PP1 activity has been shown to be required for the fusion of embryonic muscle cells.38 Considering that neurabin I is an inhibitory subunit for PP1 and an F-actin binding protein, the protein may have a key role in skeletal muscle differentiation. As such, changes in dosage of neurabin I may impair proper development of skeletal muscle.
Chimeras between normal and uniparental mouse embryos have provided an ideal system to investigate the results of genomic imprinting during prenatal and postnatal development.39–41 This is due to the fact that the selective elimination of uniparental cells has been observed in a spatial and/or temporal specific manner when imprinted genes have critical functions in embryonic development. Interestingly, skeletal muscle specific imprinting was predicted to exist based on studies using the chimeric normal/parthenogenetic embryo system42–44; selection against the parthenogenetic cells was observed in a tissue specific manner, most strikingly in tongue,44 indicating the need for tissue specific imprinted differentiation gene(s) in tongue development. The selective elimination of parthenogenetic cells from the tongue of chimeric embryos was found to occur between days 13 and 15 of development, coinciding with the beginning of myoblast fusion.44 Taking into consideration the known features of the neurabin I protein and the expression/imprinting patterns of Ppp1r9a shown in our study, neurabin I is a candidate molecule to be causal for the selective elimination of parthenogenetic cells from the tongue of chimeric embryos.
Acknowledgments
We acknowledge the technical assistance of The Centre for Applied Genomics (http://tcag.bioinfo.sickkids.on.ca/) at The Hospital for Sick Children. KN is a fellow of CIHR. SWS is an Investigator of CIHR and International Scholar of the Howard Hughes Medical Institute.
REFERENCES
Footnotes
-
↵* These two authors contributed equally to this manuscript.
-
↵† Current address: Kihara Institute for Biological Research, Graduate School of Integrated Science, Yokohama City University, Yokohama, 244-0813, Japan.
-
↵‡ Current address: Faculty of Medicine, Yamaguchi University, Yamaguchi, 753-8511, Japan.
-
This research is supported by grants from the Canadian Institutes of Health Research (CIHR) and Genome Canada.
-
Conflict of interest: none declared.
-
Data deposition: GenBank accession number, AY308065.