



Abstract
The evolving view of gut microbiota has shifted toward describing the colonic flora as a dynamic organ in continuous interaction with systemic physiologic processes. Alterations of the normal gut bacterial profile, known as dysbiosis, has been linked to a wide array of pathologies. Of particular interest is the cardiovascular–metabolic disease continuum originating from positive energy intake and high-fat diets. Accumulating evidence suggests a role for sex hormones in modulating the gut microbiome community. Such a role provides an additional layer of modulation of the early inflammatory changes culminating in negative metabolic and cardiovascular outcomes. In this review, we will shed the light on the role of sex hormones in cardiovascular dysfunction mediated by high-fat diet-induced dysbiosis, together with the possible involvement of insulin resistance and adipose tissue inflammation. Insights into novel therapeutic interventions will be discussed as well.
SIGNIFICANCE STATEMENT Increasing evidence implicates a role for dysbiosis in the cardiovascular complications of metabolic dysfunction. This minireview summarizes the available data on the sex-based differences in gut microbiota alterations associated with dietary patterns leading to metabolic impairment. A role for a differential impact of adipose tissue inflammation across sexes in mediating the cardiovascular detrimental phenotype following diet-induced dysbiosis is proposed. Better understanding of this pathway will help introduce early approaches to mitigate cardiovascular deterioration in metabolic disease.
Introduction
The gut microbiota (GM) is a complex ecosystem that can be described as a dynamic organ with an active role in human health and disease (Putignani et al., 2014). The microbial community has high plasticity and is sensitive to several stimuli including environmental, hormonal, dietary, and stress-related factors (Putignani et al., 2014). Nevertheless, diet remains one of the most vigorous modulators of GM (David et al., 2014) with Western-type calorie-dense diets driving an imbalance of microorganisms in the gut or dysbiosis (Sen et al., 2017). This is particularly relevant to the steady rise in the prevalence of metabolic disease like diabetes, obesity, and their complications, driven by increased caloric intake after the global shift to the Western diets, rich in saturated fat and refined sugars (Lutsey et al., 2008; Misra et al., 2010). As such, there has been an increasing interest in studying dysbiosis in these maladies together with the impact of its modification as a therapeutic option.
Significantly, most of the health burden associated with metabolic dysfunction is due to the high risk of cardiovascular mortality and morbidity due to ischemic heart disease, ischemic stroke, cardiac metabolic dysfunction, and heart failure (Ash-Bernal and Peterson, 2006; von Bibra et al., 2016). Of note, cardiovascular risk evoked by metabolic impairment has long been associated with a state of chronic low-grade inflammation (de Rooij et al., 2009). Indeed, under circumstances leading to dysbiosis, GM can contribute to this inflammatory state. Normally, the host health/gut bacteria interaction occurs through exposure to either bacterial components known as pathogen-associated molecular patterns, like flagella and cell wall constituents like lipopolysaccharide (LPS) (Tilg et al., 2019) or to the metabolites produced by bacterial digestion and processing of ingested food, which were shown to have several effects including modulation of the function of immune and autonomic nervous system as will be discussed below. Therefore, dysbiosis outcomes depend on the changes in bacterial phyla residing in the gut.
Interestingly, considerable sex-dependent differences were reported in inflammatory changes and cardiovascular risk associated with metabolic dysfunction. Recent literature shows that metabolic impairment in humans leads to different inflammatory profiles across sexes with increased production of proinflammatory cytokines in males (ter Horst et al., 2020). Indeed, premenopausal females are less prone to adverse cardiovascular events (Mosca et al., 2011), and varying cardiovascular profiles secondary to metabolic deterioration are observed in either sex (Gerdts and Regitz-Zagrosek, 2019). Although sex-dependent differences in metabolic-derived cardiovascular diseases (CVDs) are typically attributed to estrogen-driven alteration in insulin sensitivity, adiposity, adipocyte size and function, as well as adipose tissue (AT) susceptibility to inflammation (Ribas et al., 2010; Pradhan, 2014; Zore et al., 2018), sex-dependent differences in GM together with its vulnerability to dysbiosis add a new layer of complexity to the paradigm. GM appears to play an important role in mediating the differential patterns observed in diet-induced metabolic and cardiovascular dysfunction across sexes. Although the exact mechanism has yet to be comprehensively and systematically investigated, we attempt here to shed the light on the potential mechanisms through which dysbiosis mediates cardiovascular dysfunction in early metabolic impairment in a sex-dependent manner. We explore the sex differences in high-fat diet- (HFD) induced dysbiosis and the consequent AT inflammatory changes and cardiovascular dysfunction in the context of early metabolic deterioration. As well, we summarize some of the available evidence regarding possible therapeutic interventions to address these disorders via targeting the gut microbiome homeostasis.
Sex-Dependent Differences in Gut Microbiota in the Healthy State
The assumption of dysbiosis in disease states necessitates a fundamental knowledge of the composition and function of GM in healthy individuals. Nevertheless, a unified healthy GM profile has not been defined at any profound taxonomic resolution owing to several endogenous and exogenous factors. These include interindividual host genetic and environmental differences (Hooper et al., 2001; Abdul-Aziz et al., 2016; Rothschild et al., 2018), disparate GM growth rates, strain-level diversities, and variants within microbial genes (Huttenhower et al., 2012; Korem et al., 2015; Zeevi et al., 2019). However, high taxonomic diversity, along with high microbial gene richness and stable microbiome functions represent characteristics of a healthy GM (Huttenhower et al., 2012).
The major bacterial phyla inhabiting the human gut are Fermicutes, Bacteroidetes, Proteobacteria, Verrucomicrobia, Actinobacteria, and Fusobacteria, with Firmicutes and Bacteroidetes accounting for almost 70% of the total microbiota (Zoetendal et al., 2008; Mariat et al., 2009) and their Firmicutes/Bacteroidetes (F/B) ratio changing under situations of metabolic impairment (Turnbaugh et al., 2006). The homeostatic state in which the GM is healthy and balanced is referred to as a state of eubiosis (Iebba et al., 2016). Significantly, several studies highlighted sex-dependent variations of the GM in health and disease (Org et al., 2016; Han et al., 2017; Razavi et al., 2019). It was shown that Drosophila melanogaster strains exhibit a differential abundance of microbes across sexes irrespective of nutritional conditions (Han et al., 2017). Interestingly, similar results were also obtained in different strains of mice where the abundance of several taxa exhibited significant sex-dependent differences (Org et al., 2016). Furthermore, such differences were also evident in mice fed either normal chow or HFD, suggesting sex-by-diet interactions (Org et al., 2016).
Indeed, the impact of gonadectomy and sex hormone replacement on GM is far from settled. This is of particular interest in light of the reduced susceptibility of premenopausal women to metabolic and cardiometabolic diseases (Santos-Marcos et al., 2019) where numerous lines of evidence suggested a role for female sex hormones in enhancing the diversity of the GM (Song et al., 2020). A cross-sectional study revealed that although sex-dependent differences in GM were observed before puberty, they tended to increase after puberty with significant differences in β-diversity (interindividual dissimilarity) but not in α-diversity (intraindividual bacterial diversity) (Yuan et al., 2020). Furthermore, studies have shown a stronger impact of female gonadal hormones on GM. A study in rats showed that although the sex-dependent differences in GM community persisted after gonadectomy, the detrimental impact was more pronounced in female rats, especially when animals were overfed (Santos-Marcos et al., 2020). This is indeed in line with observations in humans demonstrating a shift in the GM profile in postmenopausal women compared with age matched men (Santos-Marcos et al., 2018). Along the same lines, the strong associations observed between sex and α-diversity in young adults, although persisted after adjusting for cardiometabolic parameters, tended to diminish after 40 years of age (de la Cuesta-Zuluaga et al., 2019), consistent with an age-related decline in the level of sex hormones. Importantly, the interaction between GM and female sex hormones appears to be bidirectional, whereby a study on rats showed that several GM-derived microRNAs were reported to modulate steroid biosynthesis and estrogen signaling (Santos-Marcos et al., 2020).
Although less clear in human studies, accumulating evidence from experiments on mice suggests that the differential diversity in GM can drive sexually dimorphic immune responses (Elderman et al., 2018; Felix et al., 2018). It was even suggested that sexual dimorphism in susceptibility to certain autoimmune disorders, like Type 1 diabetes mellitus, was mediated by GM in rodent models, and the alteration of GM at an early age may protect against genetic predisposition to autoimmune diseases (Markle et al., 2013; Yurkovetskiy et al., 2013; Candon et al., 2015). This sex-dependent dysbiosis in disease prognosis was suggested to mediate manganese-induced neurotoxicity (Chi et al., 2017). As these factors augment the complexity of the host environment-microbiota interactions, it becomes plausible that diet-induced GM alteration leading to metabolic impairment will trigger distinct inflammatory responses in either sex, culminating in disparate cardiovascular consequences.
Sex-Dependent Gut Microbiome Alterations in Early Metabolic Impairment
Early metabolic impairment has long been discussed in the literature, yet there has been no consensus on the exact definition and the diagnostic criteria. This is despite the fact that a significant proportion of the global population exhibits suboptimal metabolic health, primarily due to excessive caloric consumption and sedentary lifestyles resulting in the increased prevalence of metabolic diseases such as insulin resistance, obesity, and diabetes (Chatelier et al., 2013; Zheng et al., 2018; Jaacks et al., 2019; Frost et al., 2021). This is mirrored by an increased prevalence of metabolic dysfunction-associated cardiovascular diseases (Lakka et al., 2002). Indeed, recent research identified early stages of metabolic deterioration such as prediabetes or metabolically unhealthy normal weight as risk factors of cardiovascular disease (Stefan, 2020; Alderman, 2021). Despite the various pathologic mechanisms culminating in the emergence of these disorders, it seems that they are correlated with GM alterations referred to as dysbiosis (Qin et al., 2012; Qin et al., 2014; Allin et al., 2018). Next-generation sequencing of the gut microbiome had a major role in unfolding the involvement of GM in regulating the host metabolism. Metabolic-related dysbiosis is usually exemplified by altering the abundance of Bacteroides, Prevotella, Desulfovibrio, Lactobacillus, and Oxalobacter genera in the gut (Tyakht et al., 2013; Clemente et al., 2015; Smits et al., 2017). Nevertheless, the precise dynamics of GM involvement in metabolic diseases is not fully explored yet. Indeed, whether the disease-associated aberrant microbiota underpins disease causation or represents a secondary phenomenon after disease onset and progression has been widely debated (Bäckhed et al., 2004; Pedersen et al., 2016; Qin et al., 2010). Fig. 1 outlines the proposed framework linking dysbiosis in metabolic dysfunction with the pathogenesis of cardiovascular complications together with the interplay with sex-dependent factors as described below.

High fat diet-mediated gut dysbiosis and links to cardiovascular disease and sex-dependent factors. Significant increase in the F/B ratio occurs after the consumption of a high-fat diet. The ensuing gut microbiota dysbiosis detrimentally affects the adipose tissue and the cardiovascular system through intricate pathways. Mechanistically, gut microbiota dysbiosis enhances lipopolysaccharide production, as well as its chylomicron-mediated transport and paracellular diffusion. The latter is possible due to gut microbiota dysbiosis-mediated dysfunction of tight junctions leading to a compromised gut integrity. Locally, LPS activates, through TLR4, the proinflammatory NF-κB pathway. Systemically, the increased levels of serum LPS results in endotoxemia. Importantly, sex hormones are partly responsible for the differential modulation of these pathways in either sex as indicated. Pathways involved in gut microbiota dysbiosis are presented in red, whereas those counteracting them in black. AT; COX2, cyclooxygenase 2; ER-β; F/B; GM; HFD; IAP, intestinal alkaline phosphatase; IL; iNOS, inducible nitric oxide synthase; IR, insulin resistance; LPS; MetS, metabolic syndrome; MUC2; MyD88; NF-κB; PUFA, polyunsaturated fatty acid; TLR4; TNF-α.
Sex-Dependent Differences in Diet-Induced Dysbiosis
As diet remains the cornerstone for GM modulation in humans and animal models (David et al., 2014; Carmody et al., 2015), dysbiosis has been the focus of research on diet induced pathologies. HFD was consistently reported to provoke dysbiosis by altering the scale of the major gut phyla, increasing F/B ratio as well as an increase in Proteobacteria, which were ushered with impaired metabolic and cardiovascular function (Moreira et al., 2012; Murphy et al., 2015). Bacteroidetes is considered the most prevalent gram-negative bacteria in the gut and is essentially considered beneficial due to their capacity to modulate caloric absorption through polysaccharide metabolism (Wexler, 2007). On the other hand, Firmicutes are largely gram positive and are capable of producing various short-chain fatty acids (SCFAs) (Den Besten et al., 2013). It is generally accepted that a higher F/B ratio is usually observed in overweight and obese subjects (Ley et al., 2006; Million et al., 2012; Kasai et al., 2015), and a reduction in the F/B ratio has been associated with weight loss (Ley et al., 2006; Turnbaugh et al., 2006). However, the opposite was documented as well where HFD and diet-induced obesity were associated with decreased F/B ratio in human and animal models in both sexes (Collado et al., 2008; Schwiertz et al., 2010).
Indeed, dysbiosis severity in different situations of metabolic impairment appeared to be sex dependent (Org et al., 2016). Premenopausal women were shown to have a higher F/B ratio than postmenopausal women and men (Santos-Marcos et al., 2018). Accumulating evidence suggested that women harbor a higher F/B ratio in comparison with men, even after adjusting for body mass index (BMI) (Mueller et al., 2006; Dominianni et al., 2015). In fact, F/B ratio was found to be highly influenced by BMI and has been used as an indicator of gut dysbiosis with a higher F/B ratio indicating a more pronounced dysbiotic microbiome (Kasai et al., 2015). Another human study showed that among subjects with BMI greater than 33, men exhibited a significantly lower F/B ratio in comparison with women, whereas the opposite was observed in subjects with a BMI lower than 33 as well as in postmenopausal women (Haro et al., 2016). Sex-differential dysbiosis was also observed among lean men and women, as postmenopausal women had a similar GM signature as men, whereas obesity abolished these differences. The same study emphasized the tight correlation between sex hormones and GM diversity. Premenopausal women had higher Bifidobacterium and lower Bacteroides than men and postmenopausal women, as GM community could predict testosterone level in humans and recipient mice of human fecal microbiome (Mayneris-Perxachs et al., 2020).
In mechanistic terms, estrogen receptor-β (ERβ) has been proposed to be a modulator of sex-dependent dysbiosis. For instance, ERβ knockout female mice on isoflavone and fiber-rich feeding achieved a state of eubiosis with an increase in Bacteroidetes and a reduction in Firmicutes and Proteobacteria. However, when these mice were switched to an isocaloric low-fiber and simple-sugar rich diet, the knockout mice had more pronounced dysbiosis and reduced Bacteroidetes in favor of Proteobacteria compared with their wild-type counterparts (Menon et al., 2013), suggesting a protective effect of estrogen in diet-induced dysbiosis. Similarly, genetically obese ob/ob mice show an elevation of F/B ratio (Turnbaugh et al., 2006). Interestingly, hormonal treatment with 17β-estradiol (E2) in female mice was found to correct HFD-related dysbiosis in ob/ob and wild-type mice by increasing the heterogeneity of GM distribution and reducing the F/B ratio compared with the vehicle group (Acharya et al., 2019). Moreover, androgenization of young and adult ovariectomized female Wistar rats induced dysbiosis regardless of dietary intervention. It reduced GM diversity, elevated F/B ratio, and impaired overall metabolic function (Moreno-Indias et al., 2016).
Although human studies link dysbiosis to obesity, results from experiments on animals implicate HFD as the culprit even in absence of obesity. This was supported with results from resistin-like molecule β knockout female mice, a model that lacks the specific gene for colonic goblet cells, whereas HFD-fed mice were not obese. These mice presented with the typical HFD-related dysbiosis (Hildebrandt et al., 2009). It is noteworthy that the type of fat affects the microbiome alteration as diets rich in saturated fatty acids are thought to contribute to the development of endotoxemia by enhancing the production of LPS, whereas polyunsaturated fatty acids are suggested to exert protective effects by influencing systemic endotoxin concentrations, LPS clearance, bile acid metabolism, intestinal alkaline phosphatase activity, intestinal mucosal permeability, and microbiota composition diversity (Bellenger et al., 2019; Cândido et al., 2020). Moreover, saturated fatty acids were recently demonstrated to act as a nonmicrobial toll-like receptor 4 (TLR4) agonists, triggering myeloid differentiation primary response 88 (MyD88)-dependent or independent inflammatory pathways culminating in the activation of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and the production of inflammatory cytokines, in a similar fashion to LPS (Rocha et al., 2016). Moreover, mice fed HFD rich in omega-6 fatty acids but not omega-3 fatty acids exhibited a more pronounced metabolic endotoxemia (Kaliannan et al., 2015). Transgenic mice over-converting omega-6 to omega-3 in tissues exhibited an augmented production of intestinal alkaline phosphatase, which subsequently alters GM composition, reduces LPS production, maintains gut barrier function, and reduces endotoxemia (Kaliannan et al., 2015). Importantly, linoleic acid and α-linolenic acid-enriched HFD-fed obese mice exhibited a sex-dependent alleviation of endotoxemia and systemic and AT inflammation, which was associated with sex-dependent alterations in GM composition (Zhuang et al., 2018). This suggests that linoleic acid may provide a protective effect against metabolic endotoxemia in female mice, whereas α-linolenic acid exerts a similar effect in male mice through modulating the gut-adipose tissue axis (Zhuang et al., 2018). These variabilities highlight the complex interplay of sex and diet in shaping the microbiome community and its metabolic consequences.
Sex-Dependent Differences in Dysbiosis Consequences: Compromised Gut Integrity and Metabolic Endotoxemia
As mentioned in the previous section, the detrimental effects of consumption of HFD are partly mediated by dysbiosis evidenced by an augmented and reduced relative abundance of Proteobacteria and Bacteroidetes, respectively. Disruption of gut barrier function as well as metabolic endotoxemia ensue (Satokari, 2020). Although a sizable body of evidence consistently supports an increased F/B ratio under these circumstances, an increased LPS would not be expected given the observed reduction of the gram-negative Bacteroidetes. Paradoxically, increased plasma LPS levels were reported after HFD in animal experiments (Cani et al., 2008) and in obese humans in comparison with their lean controls (Stoll et al., 2004; Trøseid et al., 2013). Such observations can be attributed to the reduced expression of intestinal epithelial tight junction proteins leading to a compromised gut barrier in animals and humans, allowing for increased LPS transportation (Saad et al., 2016). As well, chylomicrons, lipoprotein particles mediating intestinal fat absorption, were found to facilitate LPS transport across the intestinal lumen through enterocyte-mediated absorption (Ghoshal et al., 2009), offering an additional mechanism by which HFD can contribute to increase plasma LPS levels.
Expectedly, metabolic endotoxemia will follow, whereas the transport of microbial-associated molecular patterns leads to systemic low-grade and subclinical inflammation via the activation of TLR4 (Turnbaugh et al., 2006; Cani et al., 2007; Rodriguez et al., 2020). Indeed, LPS binds to cluster of differentiation 14 and the TLR4/Myeloid differentiation factor 2 receptor complex (Kitchens and Thompson, 2005; Lu et al., 2008). TLR4 cascade was shown to mediate HFD-induced inflammation. HFD feeding led to an increased serum and fecal LPS, concurrently with increased F/B ratio in male wild-type mice. Moreover, these mice suffered from diet-induced colitis indicated by increased intestinal tumor necrosis factor α (TNFα), interleukin (IL)-1β, inducible nitric oxide synthase, cyclooxygenase 2, and phosphor-IK kinase β NF-κB expression and activity together with a compromised gut integrity and reduced expression of tight junctions protein occludin and claudin-1 (Kim et al., 2012). On the other hand, HFD-fed TLR4 knockout mice were protected from metabolic endotoxemia, as they had lower serum LPS compared with their low-fat fed counterparts. At the same time, the knockout mouse intestine did not show the same HFD-induced inflammatory consequences (Kim et al., 2012).
Sexual dimorphism in the response to HFD-induced gut barrier permeability and metabolic endotoxemia has been attributed to differential hormonal modulation of these pathologic processes. Estrogen has been shown to prevent metabolic endotoxemia and chronic low-grade inflammation, which was suggested to be a contributor to the cardiometabolic privilege of premenopausal women (Santos-Marcos et al., 2019). Indeed, 17β-estradiol-treated male, ovariectomized, and intact female mice exhibited reduced LPS production and lower susceptibility to metabolic endotoxemia and metabolic syndrome partly through upregulation of intestinal alkaline phosphatase (Kaliannan et al., 2018). On the same note, one study highlighted a protective effect of progesterone against endotoxemia, where plasma levels of LPS negatively correlated with plasma progesterone levels but positively correlated with TNF-α plasma levels in pregnant women (Zhou et al., 2019).
This differential effect of sex hormones was shown to be due to the alteration of expression and function of tight junctions and mucin production, thus regulating HFD-induced alteration of gut permeability. For example, progesterone was found to upregulate the expression of the tight junction protein occludin and inhibit NF-κB activation in LPS-stimulated Caco-2 cells (Zhou et al., 2019). Similarly, estrogens promoted gut barrier function through several mechanisms. For instance, ERβ was proposed to play a crucial role in regulating cellular differentiation in colonic tissue as ERβ−/− mice exhibit epithelial hyperproliferation and compromised integrity (Imamov et al., 2004; Wada-Hiraike et al., 2006). This suggests a homeostatic role of estrogen in the maintenance of colon integrity. Furthermore, estrogen-ERβ cascade was suggested to enhance mucin 2 (MUC2) secretion by intestinal goblet cells, which offers epithelial protection. Hence, the deletion of ERβ disrupted the colon mucin layer in female mice (Wada-Hiraike et al., 2006; Diebel et al., 2015). Likewise, female mice during proestrus stage, characterized by high estrogen levels, are protected against intestinal injury in comparison with their male counterparts and female mice in the diestrus stage that is characterized by low circulating estrogen (Homma et al., 2005; Sheth et al., 2010). This can be attributed to the lower mucus thickness observed in the colon of diestrus female mice and male mice compared with the proestrus ones (Elderman et al., 2017). Apart from the induction of MUC2 production, estrogen is also found to upregulate the expression of tight junctions in both male and ovariectomized female rodents as well as in vitro monolayer cultures (Homma et al., 2005; Braniste et al., 2009; Looijer-van Langen et al., 2011). Consistent with these observations, the differential effects of estrogen supplementation on metabolic health observed in pre- and postmenopausal women suggest that the loss of ovarian function may profoundly alter estrogen signaling (Hulley et al., 1998; Rossouw et al., 2002). Indeed, it was shown that ovariectomized mice exhibit temporal and regional changes in gastrointestinal permeability due to disruption of tight junctions (Collins et al., 2017). Intriguingly, it was shown in some studies that TLR4 level in female macrophages were higher after ovariectomy (Rettew et al., 2009). Although males tend to have higher metabolic endotoxemia, orchiectomized mice were more susceptible to endotoxemia, and isolated macrophages presented higher TLR4 level than intact males. At the same time, testosterone treatment attenuated these events, suggesting an immunosuppressive effect of testosterone (Rettew et al., 2008). These findings emphasize the complexity of the effect of sex hormones on dysbiosis outcomes. Nevertheless, female sex hormones in animal models consistently exhibited a protective effect on gut integrity and the resulting metabolic endotoxemia, mainly by upregulating intestinal tight junction proteins.
Sex-Dependent Differences in Dysbiosis-Induced Insulin Resistance and Adipose Inflammation
Diet-induced metabolic derangement and its correlated cardiovascular dysfunction have been suggested to be an outcome of early adipose inflammation and insulin resistance, even in the absence of explicit hyperglycemia (Shah et al., 2008; Nishimura et al., 2009; Gollasch, 2017; Elkhatib et al., 2019; Rafeh et al., 2020). As previously demonstrated, dysbiosis and altered gut permeability precede the emergence of metabolic syndrome (Martínez-Oca et al., 2020). Since HFD consumption increased intestinal permeability by impairing the function of tight junctions (Cani et al., 2008), intestinal hyperpermeability and GM dysbiosis were suggested to further the inflammatory phenotype in AT and to increase the risk of cardiovascular diseases (Serino et al., 2012; Kallio et al., 2015; Clemente-Postigo et al., 2019; Gasmi et al., 2021). A great body of research attempted to establish an understanding of the gut-adipose axis in metabolic dysfunction (Poggi et al., 2007; Samuel et al., 2008; Serino et al., 2012). Accordingly, white adipose tissue (WAT) was identified as a major target of GM. Not only were GM fermentation products shown to regulate WAT energy balance (Samuel et al., 2008), but also GM regulated fat deposition in AT as well as insulin resistance (Bäckhed et al., 2004; Bäckhed et al., 2007; Velagapudi et al., 2010). Such observations are of relevance given the contribution of WAT to the development of metabolic inflammation leading to insulin resistance and cardiovascular dysfunction (Bouloumié et al., 2005; Hotamisligil, 2006; Bouloumié et al., 2008). In HFD-fed diabetic mice, dysbiosis increased the stromal vascular fraction in WAT, in particular macrophages, lymphocytes, and preadipocytes (Serino et al., 2012). Additionally, LPS migration into the circulation has been suggested to be a contributing factor to the onset of AT inflammation, insulin resistance, obesity, and diabetes (Cani et al., 2007; Cani et al., 2008; Hersoug et al., 2016). Interestingly, HFD consumption had a similar effect to LPS subcutaneous infusion on elevating serum LPS and promoting AT inflammation in male mice (Cani et al., 2007). Significantly, HFD-fed TLR4 knockout mice did not have increased proinflammatory cytokines in the isolated epidydimal AT depot, whereas wild-type mice were hyperinsulinemic and exhibited a proinflammatory response in the epidydimal WAT manifested by increased TNFα, IL-1β, and IL-6, and macrophage infiltration (Kim et al., 2012). Moreover, TLR4 knockout male mice had an increased insulin sensitivity in subcutaneous and epidydimal WAT even in presence of HFD feeding (Poggi et al., 2007). Consistently, HFD-fed male mice showed an improved metabolic status after eight weeks of antibiotic treatment, as serum LPS, insulin, and fasting glucose were reduced. The epidydimal WAT in the treated group had lower TLR4 activation, Jun NH2-terminal kinase inactivation, lower nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor α degradation, and inhibition of insulin receptor substrate 1 Ser307 phosphorylation. Additionally, the treatment group had improved insulin signaling with increased protein kinase B (Akt) phosphorylation and reduced macrophage infiltration. It is noteworthy that HFD-fed germ-free mouse models were not vulnerable to HFD-induced insulin resistance (Fleissner et al., 2010; Rabot et al., 2010).
As for sex-based differences, a recent study reported that HFD induced weight gain and insulin resistance in males but not in female mice with differences in gut microbiome recorded at baseline (Peng et al., 2020). Moreover, E2 treatment in intact female mice improved HFD-induced weight gain, glucose intolerance, and insulin resistance possibly as a result of the downregulation of lipogenic genes, such as sterol response element binding protein-1 (SREBP-1) and leptin and resistin genes expression in WAT (Bryzgalova et al., 2008). Moreover, E2 intervention in ovariectomized female mice prevented HFD-induced obesity (Bless et al., 2014). Administration of E2 in ovariectomized female rats increased leptin sensitivity and led to preferential subcutaneous adiposity, resembling the intact female littermates (Clegg et al., 2006). Notably, subcutaneous fat pads were suggested to be more sensitive to insulin than visceral fat (Chang et al., 2018). This might be one of the many protective roles of estrogens against metabolic impairment related to AT dysfunction, as premenopausal women have more subcutaneous AT than men and postmenopausal women (Lemieux et al., 1993). Furthermore, E2 was proposed to ubiquitinate and degrade HIF1-α, eventually reducing adipose inflammation and its subsequent metabolic derangements (Kim et al., 2014). Typically, insulin resistance is associated with hypertrophic adipose expansion leading to increased tissue hypoxia and elevated hypoxia inducible factor 1-α (HIF1-α) expression, which was suggested to contribute the inflammatory cascade (Lumeng et al., 2007; Palmer and Clegg, 2014; Wensveen et al., 2015).
Since adiposity patterns, insulin sensitivity, LPS levels, TLR activation, hypoxia, and AT inflammation were all ameliorated in the presence of estrogen, its enhancement of AT metabolic state via GM modulation can be one of the protective roles female sex hormones possess against cardiometabolic dysfunction. Interestingly, HFD-fed androgen receptor knockout male mice had increased obesity, visceral adiposity and adipocyte hypertrophy, glucose intolerance, and insulin resistance compared with control males and HFD-fed females. These derangements were linked to dysbiosis as antibiotic treatment corrected the dysfunction (Harada et al., 2020). These observations further emphasize the complexity and the importance of sex hormone contribution to the GM-cardiometabolic interaction, particularly the female sex hormones playing a modulatory role on several intermediary factors of cardiometabolic insults as outlined above. Figure 2 depicts the different pathways triggered by GM, leading to WAT inflammation as well as their modulation by sex hormones.

Detrimental effects of dysbiosis on adipose tissue homeostasis. Gut microbiota dysbiosis amplifies high-fat diet-mediated adipose tissue dysfunction through increasing energy absorption, weight gain, and adiposity. The subsequent development of insulin resistance and adipose tissue inflammation leads to the development of the metabolic syndrome linked to the emergence of cardiovascular diseases. Testosterone and estrogen inhibit TLR4 signaling. Estrogen decreases the level of circulating LPS, enhances leptin sensitivity, decreases insulin resistance, and limits consequences of hypoxia by induction of the proteasomal degradation of HIF-1α. Several therapeutic interventions such as antibiotic treatment and fecal microbiota transfer also positively modulate the depicted pathways. Pathways implicated in gut microbiota dysbiosis are presented in red, whereas those counteracting them are presented in black. AT; ATB, antibiotic; CVD; FMT; GM; HFD; HIF-1α, hypoxia-inducible factor 1α; IL; MyD88; NF-κB, nuclear factor κ B; TLR4; TNF-α.
Nonetheless, it is prudent to mention that accumulating evidence recognize sex hormone independent differences in metabolism and response to metabolic challenge between males and females (Manwani et al., 2015). Differences in consequences of the genetic disparity between the XX and XY chromosome combination extend well beyond sex hormone production to comprise variable levels of X-linked gene imprinting and expression, in addition to a considerable dimorphism in the complement of noncoding RNA molecules production (Link et al., 2013). The study of the metabolic sequalae of such differences was hampered for a long time by the lack of ability of dissociating the concurrence of the XX or the XY combination with the corresponding gonadal hormone production. Evidence from transgenic mouse models expressing different combinations of sex chromosomes in presence and absence of the corresponding gonads showed a remarkable impact of the XX chromosome combination on preferential subcutaneous AT distribution regardless of the gonadal hormone status (Chen et al., 2012). However, the increased subcutaneous adiposity in this model was associated with increased insulin resistance, thus emphasizing the protective effect of estrogen in this regard. More recently, genetic association studies were conducted on humans to detect the effect of the X chromosome on metabolic and cardiovascular disease but had conflicting results. Although some studies found differential associations with insulin resistance, atherosclerosis, and coronary artery disease between sexes (Tukiainen et al., 2014; Traglia et al., 2017), others failed to detect these differences and attributed the observations to gonadal hormones (Manwani et al., 2015). However, to our knowledge, the interaction between sex chromosomes and dysbiosis did not receive much attention and warrants detailed investigation in the future.
Sex-Dependent Differences in Alteration of Short-Chain Fatty Acids Production and Consequent Metabolic Dysfunction
Another important component of HFD-induced dysbiosis is the decline in SCFA generation, mainly acetate, propionate, and butyrate, which were correlated with numerous metabolic and cardiovascular disorders (Canfora et al., 2015; Chambers et al., 2018). Despite the extensive research on the role of SCFA in the host wellbeing, the exact role of SCFAs in regulating metabolic function remains debatable. For instance, some evidence suggested a negative correlation of fecal SCFAs levels with the host’s metabolic function (Teixeira et al., 2013; de la Cuesta-Zuluaga et al., 2018) with studies showing that SCFA levels were higher As for acetate and propionate, it is suggested that they offset LPS induced endotoxemia by lowering TNF-α and NF-κB production as shown in human neutrophils and macrophages in vitro experiments (Canfora et al., 2015). These two SCFAs were shown to be effective in reducing TNFα production in LPS-activated neutrophils, while repressing the activity of NF-κB receptor in a human colon adenocarcinoma cell line (Tedelind et al., 2007). Consistently, in vitro treatment of human omental and subcutaneous adipocytes with propionate, reduced mRNA expression of the proinflammatory factor resistin, and stimulated leptin mRNA expression (Al‐Lahham et al., 2010). Another study on human omental AT supported the previous findings, as propionate treatment declined both mRNA and protein levels of proinflammatory cytokines like IL-4 and TNF-α (Al‐Lahham et al., 2012). Additionally, animal models reared on HFD and treated with propionate had an improved insulin sensitivity, glucose tolerance, thermogenesis, and mitochondrial function, besides an improved metabolic state in brown adipose tissue, liver, and muscles (Liang and Ward, 2006; Brial et al., 2018). In humans, rectal administration of sodium acetate reduced serum TNF-α in obese females (Freeland and Wolever, 2010). Another trial on acute intravenous infusion of acetate in women with hyperinsulinemia and overweight improved serum peptide YY, glucagon-like peptide 1 and reduced circulating TNFα and ghrelin (Freeland and Wolever, 2010). Pathways modulated by SCFAs are summarized in Figure 3.

Short-chain fatty acids regulate lipogenic, inflammatory, and neuronal pathways that are dysregulated in states of metabolic dysfunction. The three major short-chain fatty acids are acetate, propionate, and butyrate. They improve cardiometabolic health through several pathways as indicated, thus counteracting HFD-induced metabolic dysfunction. Akt; BP; FIAF, fasting-induced adipose factor; FMT; GPR; HFD; IL; LPL, lipoprotein lipase; LPS; MAPK, mitogen-activated protein kinase; NF-κB; PKC; protein kinase C; PLC; PTEN, phosphatase and tensin homolog; SCFA; SREBP-1; TNF-α; UCP-1, uncoupling protein 1.
Importantly, recent literature reveals that interventions targeting GM have a differential impact on SCFA generation across sexes. For instance, ciprofloxacin-metronidazole treatment reduced SCFA production only in female mice consistent with a reduction in the relative abundance of Firmicutes (Gao et al., 2019). Alternatively, prebiotic supplementation was shown to increase fecal butyrate output only in male but not female rats (Shastri et al., 2015). A similar study in humans demonstrated a differential effect whereby beta-glucan supplementation led to an increased butyrate production in female subjects but not in males (Trimigno et al., 2017). On the other hand, SCFAs were proposed to promote the storage of triglycerides through the activation of lipogenic hepatic enzymes including SREBP, which show a positive differential expression in women (Bäckhed et al., 2004; Jiang et al., 2016). This is suggested to reflect in an increased microbiota-dependent lipid storage and obesity risk in women. SCFAs also are known to suppress the fasting-induced adipocyte factor, an inhibitor of lipoprotein lipase (Bäckhed et al., 2004; Khan et al., 2016). This increased lipoprotein lipase activity may lead to microbiota-dependent augmentation in fat storage, which may contribute in part to sex differences in body composition (Bäckhed et al., 2004). Henceforth, sex-differential response to GPR41 may contribute to microbiota-associated body weight sexual dimorphism (Inoue et al., 2014). This is particularly important as male but not female GPR41 knockout mice exhibited an increased body fat mass and a decreased energy expenditure (Bellahcene et al., 2013). Moreover, further sex-specific interactions with SCFAs were reported where butyrate was shown to increase estrogen secretion in granulosa cell culture models (Lu et al., 2017). Indeed, conclusions about the involvement of SCFAs in cardiometabolic derangements in sex-dependent manner are hard to be drawn, and further studies are highly needed for more solid evidence.
Sex-Dependent Impact of Gut Microbiome on Metabolically Induced Cardiovascular Dysfunction
Although traditional cardiovascular risk factors appear to be related to the development of cardiovascular disease in either sex, research has long recognized significant complexity in their differential roles and relative weights (Njølstad et al., 1996). For instance, analysis of a large case-control study showed that the impact of diabetes and hypertension was more pronounced on the development of myocardial infarction in women than in men (Anand et al., 2008). Yet, the impact of these two factors appeared to differ by age, being stronger in younger men, leading to an age difference of cardiovascular disease onset by about nine years. This further implicates the role of sex hormones in the observed protective effect in premenopausal females, as the incidence of both hypertension (Lima et al., 2012) and diabetes (Heianza et al., 2013) in postmenopausal females appear to exceed that in men. Interestingly, it is well recognized that both disorders have strong mechanistic links to adipose tissue dysfunction, particularly perivascular adipose tissue (PVAT), observed in metabolic impairment (Saxton et al., 2019). Significantly, female-specific risk factors for cardiovascular disease such as polycystic ovary syndrome and preeclampsia appear to have a strong metabolic impairment component carrying the hallmarks of a dysfunctional adipose tissue (Huda et al., 2017; Leon et al., 2019; Osibogun et al., 2020). In the below sections, we examined the impact of sex-based differences in the interconnection among dysbiosis, metabolic impairment, and adipose inflammation on incidence of cardiovascular disease.
Dysbiosis and Cardiovascular Dysfunction
HFD is known to induce cardiovascular dysfunction (Martins et al., 2015; Aghajani et al., 2017). Since HFD stimulates dysbiosis, the microbiome-cardiovascular axis was extensively studied, and dysbiosis was linked to several diseases such as hypertension (HTN), atherosclerosis, and heart failure (HF) among others (Tang and Hazen, 2017; Taylor and Takemiya, 2017; Kappel and Federici, 2019; Razavi et al., 2019). For instance, atherosclerotic plaques were found to contain bacterial DNA, and these bacterial taxa were also present in the gut of the same individuals (Ott et al., 2006; Koren et al., 2011), proposing a possible role of microbial communities in plaque instability and the subsequent adverse effects (Koren et al., 2011). In patients with HF, both metabolites and gut flora print were significantly determinantal compared with healthy subjects and were even worse in patients with decompensated heart failure (Hayashi et al., 2018). Significantly, gut dysfunction involving disturbances in intestinal motility and villi absorption, in addition to an impaired tissue perfusion and edema, was also observed in HF patients (Krack et al., 2005; Sandek et al., 2012). Undeniably, the HFD-induced impairment of gut integrity and gut hyperpermeability were linked to the aforementioned cardiovascular insults (Lewis and Taylor, 2020).
Gut microbial signature in HTN was heavily investigated in the last few decades (Mell et al., 2015; Kim et al., 2018). Interestingly, fecal microbial transplantation from hypertensive patients to germ-free mice induced HTN in these mice (Li et al., 2017). Moreover, these results were also observed in germ-free rats receiving GM from spontaneously hypertensive rats (Shi et al., 2021). However, the impact of GM appears to be complex as germ-free rats demonstrated a reversal of poor vascular contractility and reduced blood pressure control upon acquisition of normal GM (Joe et al., 2020). Indeed, a decrease in microbial richness and diversity in prehypertensive and hypertensive human subjects were recorded as well (Li et al., 2017). Additionally, spontaneously hypertensive and chronic angiotensin-II-induced hypertensive rat models presented dysbiosis manifested by an increase in F/B ratio compared with the normotensive controls (Yang et al., 2015; Santisteban et al., 2017). Moreover, high-salt diet was shown to deplete a strain of Lactobacillus; however, treating these mice with this strain attenuated salt-sensitive hypertension (Wilck et al., 2017). Furthermore, HFD-induced dysbiosis was shown to have a role in the development of the obstructive sleep apnea-induced HTN (Durgan et al., 2016). HFD-induced dysbiosis was also correlated with increased serum LPS-binding protein, IL-6, endothelial dysfunction, arterial stiffness, aortic phosphorylated NF-κB, and NADPH oxidase in PVAT, leading to a positive oxidative state. Interestingly, all these insults were attenuated with antibiotic treatment (Battson et al., 2018). Not only had been NADPH oxidase (NOX)-related reactive oxygen species shown to be detrimental in CVDs (Brandes et al., 2010), PVAT inflammatory and oxidative changes were consistently reported to contribute to vascular and cardiac autonomic dysfunction in HFD-fed rats even prior to the development of overt metabolic impairment (Al-Assi et al., 2018; Elkhatib et al., 2019; Rafeh et al., 2020), implicating the GM-metabolic-AT-CVD axis, which will be discussed later.
SCFAs may also contribute to the reduction of systemic blood pressure and serum cholesterol levels (Den Besten et al., 2013; Mariño et al., 2017). High-fiber diet, diet supplemented with SCFA, or parenteral injection of SCFA improved cardiometabolic health in several murine models by reducing blood pressure and cardiac fibrosis (Brial et al., 2018). It was proposed that the role of SCFA in blood pressure (BP) regulation might be mediated by the activation of GPR41 in the vascular endothelium (Jonsson and Bäckhed, 2017). As well, SCFAs were found to hold strong vasorelaxant properties (Poll et al., 2020). Interestingly, they were also discovered to be ligands for olfactory receptor 78, a G protein-coupled receptor expressed in the vasculature, which plays an important role in vasoregulation and renin release and is activated mainly by acetate and propionate. Olfactory receptor 78 knockout mice had basal hypotension and low serum renin level, possibly indicative of the opposing response of GPR41 to SCFAs (Pluznick et al., 2013; Pluznick, 2014).
Importantly, SCFAs may also participate in regulating the sympathetic tone. Actually, in GPR41 knockout male mice, propionate was found to be a potent activator of sympathetic ganglia through Gβγ-PLCβ-MAPK pathway rather than cAMP synthesis inhibition (Kimura et al., 2011). Interestingly, propionate was able to trigger epinephrine secretion in sympathetic neurons. On the other hand, wild-type mice did not have GPR41 expression in the mesenteric fat pad and only presented GPR43 whose activation seemed to increase leptin production. Henceforth, it was suggested that GPR43 mediate sympathetic stimulation by adipocytes activation and leptin overexpression (Kimura et al., 2011). A recent study on Wistar-Kyoto and spontaneously hypertensive male rats revealed the strong association between dysbiosis and sympathetic activation through the induction of inflammation and oxidative stress in the brain. Not only blood pressure was corrected after fecal microbial transplantation from Wistar-Kyoto to spontaneously hypertensive rats, but also inflammation and oxidative stress in the paraventricular nucleus were improved. However, fecal microbial transplantation in the opposite direction deteriorated the inflammatory, oxidative, and blood pressure state of the Wistar-Kyoto rats simultaneously with poor gut integrity and increased colonic TNF-α and circulating LPS, which were attenuated in the former transplant. Worth mentioning, spontaneously hypertensive rats had a lower butyrate receptor expression in the hypothalamus, alongside a higher Th17 cells and macrophage infiltration in the paraventricular nucleus (Toral et al., 2019). Moreover, LPS infusion was found to induce HTN in normotensive rats, by provoking neuroinflammation in the rostral ventrolateral medulla, which is considered an important part in intensifying the sympathetic stream to the blood vessels (Wu et al., 2012). Interestingly, another study on the same model indicated a similar pattern of dysbiosis; increased F/B ratio, in addition to a notable reduction in acetate, increased gut sympathetic outflow, elevated blood pressure, impaired gut integrity, impaired endothelial-dependent relaxation to acetylcholine, and NOX overactivation. Treating spontaneously hypertensive rats with losartan, an angiotensin 2 receptor (AT2-R) antagonist, preserved gut integrity and enhanced functionality and immune response in the vasculature, improving acetylcholine-induced relaxation and increased Tregs infiltration (Santisteban et al., 2017; Robles‐Vera et al., 2020). Indeed, these lines of evidence support the involvement of dysbiosis in inducing diet-related HTN and sympathetic overactivation, alongside with neuroinflammation and oxidation, which is proposed to be corrected by SCFAs. Figure 4 depicts the role of GM in the interaction between the dysbiosis, cardiometabolic dysfunction, and neuroinflammation.

Gut microbiota dysbiosis in the metabolic dysfunction-neuroinflammation-cardiovascular disease continuum. Gut microbiota increases the F/B ratio resulting in an oxidative stress in the brain leading to neuroinflammation and sympathetic overactivation. The latter consequently increases blood pressure and predisposes to the development of hypertension. Gut microbiota dysbiosis also leads to endotoxemia and increases Th17 and macrophage brain infiltration, both leading to arterial hypertension. Gut microbiota dysbiosis also detrimentally accelerates adipose tissue dysfunction, leading to an increased production of leptin, which further augments the activation of the sympathetic system. Additionally, endotoxemia inhibits the rather beneficial accumulation of Treg cells in the brain, which further augments inflammation. Estrogen inhibits the development of hypertension through an eNOS-mediated vasodilatory effect and through decreasing serum levels of AngII. Progesterone as well as fecal microbiota transfer inhibits neuroinflammation and its downstream consequences. The antagonism of AT2-R also reverses immune cell profile alterations mediated by gut microbiota dysbiosis. Pathways involved in gut microbiota dysbiosis are presented in red, whereas those counteracting them in black. AngII, angiotensin II; AT; AT2-R; BP; eNOS, endothelial nitric oxide synthase; F/B; FMT; GM; LPS; Th17, T helper cell 17; Treg.
On another note, accumulating evidence highlighted the role of dysbiosis in augmenting the production of the bacterial metabolite trimethylamine-N-oxide (TMAO), which is suggested to be an indicator of CVDs (Moludi et al., 2020). TMAO is a plasma metabolite formed through a metaorganismal pathway, and its level is depending on dietary intake mainly from animal protein sources like red meat, egg yolk, and seafood, which are abundant in choline, phosphatidylcholine, and L-carnitine (Anders et al., 2013; Tang et al., 2015). These compounds are initially metabolized by GM to form trimethylamine (TMA) and then converted by the host liver enzyme flavinmonooxygenase 3 (FMO3) to form TMAO (Bennett et al., 2013). Noteworthy, FMO3 knockdown in female mice protected from diet-induced obesity and reduced hypertrophy and adiposity in WAT while improving total metabolic health (Schugar et al., 2017). Increased TMAO has been anticipated to induce insulin resistance and adipose inflammation in mice as well as increasing the risk for type 2 diabetes in human subjects (Tang et al., 2017). Circulating TMAO was linked to elevated vascular inflammation through the incitement of proinflammatory cytokines expression and leukocytes recruitment (Seldin et al., 2016). Also, dietary supplementation of choline in mice increased TMAO levels, macrophage foam cell formation, and atherosclerosis incidence (Wang et al., 2011). Moreover, it prompted platelet hyperactivity (Trip et al., 1990; Marcucci et al., 2014) and enhanced thrombosis (Zhu et al., 2016), which are considered major risk factors for developing CVDs. Indeed, serum TMAO level was tightly correlated with atherosclerosis (Tang and Hazen, 2017). In the same context, a human study revealed that subjects with higher serum TMAO levels had a twofold risk increase for developing major cardiovascular events compared with subjects with low TMAO (Tang et al., 2013). TMAO levels had also been suggested to be an accurate indicator for heart failure diagnosis (Tang et al., 2014). It was found to fuel endothelial dysfunction as well by upregulating vascular adhesion molecule-1, monocyte attraction, and NF-κB activation (Ma et al., 2017).
As stated before, HFD induces AT dysfunction, which predispose to metabolic and cardiovascular disorders. The AT surrounding the vascular bed, referred to as PVAT, which has been identified as a crucial component of the vascular regulatory machinery. It is worth mentioning that PVAT has been suggested to be one of the most sensitive AT depots to positive energy intake and the first to undergo negative remodeling including hypertrophy, inflammation, and hypoxia, which were observed in early metabolic impairment (Elkhatib et al., 2019; AlZaim et al., 2020). Moreover, resistance arterioles from obese mice showed a PVAT-dependent impairment in insulin/Akt-mediated vasodilatation due to reduced adiponectin and AMPK downstream effects, which was restored with Jun NH2-terminal kinase inhibition (Meijer et al., 2013). HFD induced PVAT dysfunction might be mediated via sympathetic overactivation and insulin resistance that have been linked to a wide range of subclinical cardiovascular insults such as endothelial dysfunction and cardiac autonomic neuropathy (Britton and Fox, 2011; Bulloch and Daly, 2014; Greenstein et al., 2009; Akoumianakis et al., 2017; Alaaeddine et al., 2019; AlZaim et al., 2020; Bakkar et al., 2020; Rafeh et al., 2020). As such, it becomes plausible that PVAT inflammation might mediate the effect of HFD-induced dysbiosis on early metabolic impairment and cardiovascular dysfunction. This hypothesis is presented in Figure. 5. However, limited studies have explored the association between PVAT and HFD-induced dysbiosis. One recent study examined the role of PVAT FMO3 in response to direct TMA stimulation in tone regulation in excised aortas from male rats. TMA exerted a contractile effect through activating L-type voltage-gated calcium channels that was found to be dependent on endothelium rather than PVAT, suggesting that TMAO and TMA modulate vascular tone by a direct effect of vascular smooth muscle cells (Restini et al., 2021). However, to our knowledge no studies were conducted to investigate the impact of dysbiosis or SCFAs on PVAT modulation and thus the HFD-dysbiosis-PVAT axis remains elusive.

Perivascular adipose tissue dysfunction: Novel mechanisms of gut microbiota dysbiosis-mediated cardiovascular derangements. A healthy perivascular adipose tissue secrets adiponectin, which elicits an AMPK-mediated anticontractile effect. The consumption of a high-fat diet causes sympathetic overactivation and insulin resistance, leading to perivascular adipose tissue expansion, adipocyte hypertrophy, and adipose tissue inflammation, thus jeopardizing the anticontractile activity of perivascular adipose tissue. Importantly, the consumption of a high-fat diet causes gut microbiota dysbiosis, which enhances NOX-mediated production of ROS, a pathway that is augmented after ovariectomy. Additionally, gut microbiota dysbiosis increases aortic NF-κB signaling, leading to arterial stiffness. Microbial metabolism products including TMA and TMAO participate in gut microbiota dysbiosis-caused perivascular adipose tissue dysfunction. TMA, through its activity on VSMCs induces L-type voltage-gated calcium channels, which counteracts perivascular adipose tissue-mediated anti-contractile effect. AMPK; GM; HFD; NOX; PVAT; ROS; TMA; TMAO; VCAM-1; VSMCs, vascular smooth muscle cells.
Sex-Dependent Impact of Dysbiosis on Cardiovascular Dysfunction
Sex-dependent cardiovascular risk and pathology are well documented in the literature, as men have a higher absolute risk compared with premenopausal women, a difference that diminishes after menopause, indicating the important role of sex hormones in CVDs (Kim and Reaven, 2013; Pei et al., 2017; WHO, 2017; Chella Krishnan et al., 2018). Although metabolic disorders and cardiovascular diseases have long been intertwined, early metabolic impairment has been the focus of interest as it imparts predisposition to inevitable CVDs. This effect is mainly mediated by subclinical events such as metabolic endotoxemia, AT inflammation, and insulin resistance (Heilbronn and Campbell, 2008; Shah et al., 2008; Nishimura et al., 2009; Kallio et al., 2015; Wensveen et al., 2015). Interestingly, GM has been identified as a major driver of these anomalies. Since sex-dependent differences exist in dysbiosis, the differential GM effect is expected to be extrapolated to CVDs. As discussed previously, estrogens attenuate HFD-induced gut hyperpermeability and LPS transport either through leaky membranes or chylomicrons together with the consequent metabolic endotoxemia, Th17 cell activation, and Tregs inhibition (Cani et al., 2007), which were found to be higher in men. As stated previously, LPS activates TLR4 on target tissues including AT and macrophages, triggering proinflammatory cascade and activating NF-κB. AT overactivation and hypertrophied expansion due to dysbiosis was linked to increased leptin production, which in turn will activate sympathetic outflow. In parallel, LPS-induced neuroinflammation triggers sympathetic firing. The resultant sympathetic overactivation and insulin resistance, which are expected to be higher in males, will lead to early inflammation and negative remodeling of PVAT, precipitating a wide range of subclinical cardiovascular insults (Greenstein et al., 2009; Britton and Fox, 2011; Bulloch and Daly, 2014; Akoumianakis et al., 2017; Khatib et al., 2018; Alaaeddine et al., 2019; Bakkar et al., 2020). Since estrogen holds a protective effect against hypoxia in AT, possibly estrogen will block dysbiosis-mediated dysfunction cardiovascular dysfunction by interfering with PVAT inflammation. Additionally, progesterone was found to have an anti-inflammatory effect against LPS mediated neuroinflammation (Lei et al., 2014).
The evidence regarding sexual dimorphism in HFD-induced PVAT remodeling is scarce; however, females tend to have a more functional PVAT compared with males. In this regard, ovariectomy in murine models instigated endothelial and PVAT dysfunction mediated by increased reactive oxygen species when compared with their sham littermate (Wang et al., 2014; Taylor and Sullivan, 2016). Additionally, an enhanced anticontractile role of PVAT was observed in female pigs and was attributed to a higher sensitivity of adiponectin receptor in coronary artery (Ahmad et al., 2017). Taken together, these observations suggest a possible role of PVAT in mediating a sex-dependent cardiovascular impact of dysbiosis in early metabolic dysfunction.
On another note, the protective effect of estrogen on HTN development is mainly through inducing endothelial nitric oxide synthase mediated vasorelaxation (Sobrino et al., 2017; Bucci et al., 2002), while at the same time inhibiting vasoconstricting agents such as angiotensin-II (Schunkert et al., 1997). However, mounting evidence suggests the role of immune responses in mediating sex-dependent GM-HTN axis. For instance, dysbiosis was linked to an increased activity of Th17 cells, which had a role in initiating arterial hypertension (Guzik et al., 2007; Ivanov et al., 2009; Wenzel et al., 2016). In this regard, hypertensive male rats had higher Th17 activity compared with females (Gillis and Sullivan, 2016).
Therapeutic Interventions for Cardiometabolic Consequences of Dysbiosis
Bidirectional interactions between GM and cardiovascular drugs have been reported for quite some time. Indeed, not only has the gut bacterial community been implicated in altering the pharmacokinetics of some cardiovascular drugs, but treatment with certain drug classes has also been associated with favorable changes in GM populations. For instance, certain phyla of gut bacteria were shown to metabolize digoxin and amlodipine, reducing their availability at target tissues, whereas others were proposed to decrease the absorption of simvastatin and captopril (Tuteja and Ferguson, 2019). In parallel, one human study showed that the low-density lipoprotein cholesterol lowering effect of a 4–8-week rosuvastatin treatment was associated with a change in the abundance of Firmicutes (Liu et al., 2018). As for animal studies, atorvastatin therapy appeared to reverse HFD-induced dysbiosis in male rats (Khan et al., 2018). Similarly, captopril treatment reduced dysbiosis and improved gut permeability associated with hypertension in spontaneously hypertensive rats (Santisteban et al., 2017). However, none of these interventions has been examined systematically, and the underlying mechanisms remain unclear. From a different perspective, tailored pharmacological interventions targeting GM with the purpose of imparting protective cardiovascular research have been proposed. Indeed, as bacterial metabolic reactions have been thoroughly recognized, selective approaches could be designed to modify harmful metabolite production. As such, small molecule inhibitors of TMAO synthesis were designed and proposed to treat atherosclerosis (Wang et al., 2015). Moreover, nanoparticle-based approaches were proposed either to deliver useful bacterial species associated with increased SCFA production or reduced LPS, or to scavenge TMAO and proinflammatory cytokines (Kazemian et al., 2020). Nevertheless, all these interventions remain in early stages, and the current viable options for prevention of detrimental outcomes of gut bacterial alteration remain related to direct manipulation of bacterial population using probiotics, antibiotics, fecal microbial transplantation, or using bacterial metabolites such as SCFAs as described below. The impact of these interventions on dysbiosis triggered pathways are demonstrated throughout Figures 1–5.
Probiotics
Probiotics are nonpathogenic strains of bacteria, usually belonging to Lactobacilli and Bifidobacteria, which have been used to reset microbiome dysbiosis (Holzapfel and Schillinger, 2002; Isolauri et al., 2004; Williams, 2010). Some clinical trials attempted to explore probiotics as a potential intervention with the symptoms in some neurologic and psychologic diseases such as amiotrophic lateral sclerosis and schizophrenia (Severance et al., 2017; Mazzini et al., 2018), whereas others focused on using different strains of probiotic bacteria as a potential therapy and early preventive technique for cardiovascular and metabolic disorders. However, these trials have had controversial findings. On one hand, several studies showed that probiotic administration improved cardiometabolic and inflammatory parameters not only in metabolically impaired but also in borderline individuals. For instance, a double-blind placebo-controlled trial showed that daily ingestion of Lactobacillus plantarum in hypercholesteremic individuals for 12 weeks significantly improved blood pressure, reduced serum total cholesterol, LDL, and triglycerides, while increasing high-density lipoprotein (HDL) levels (Costabile et al., 2017). The same strain was used for 6 weeks in subjects who smoked and had similar findings, reducing CV risk factors (Naruszewicz et al., 2002). In postmenopausal women with metabolic syndrome, supplementation with the same strain for 90 days decreased blood glucose and homocysteine (Barreto et al., 2014). On the same note, Lactobacillus plantarum 299v supplementation for 6 weeks in men with stable coronary artery disease significantly improved endothelium dependent vasodilatation, induced some changes in GM by enriching Lactobacillus genus, decreased plasma propionate, leptin and IL-8 and 12, without changing blood glucose, lipid profile, and body weight (Malik et al., 2018). Another strain of bacteria, Bifidobacterium longum BB536, exhibited beneficial effects after 12 weeks of blinded controlled intervention of food supplement intake containing red yeast extract, niacin, and coenzyme Q10 on individuals with a low score of CV risk. The results showed in improved levels of atherogenic lipid profile (Ruscica et al., 2019). Men with mild hypercholesteremia were treated with isoflavone-supplemented soy product fermented with Enterococcus faecium CRL 183 and Lactobacillus helveticus 416 for 42 days and showed an improved serum lipid profile, but neither CRP nor fibrinogen (Cavallini et al., 2016). Another study examined the role of Bifidobacterium lactis in patients with metabolic syndrome. The organism was supplemented in fermented milk and given for 45 days. Treated subjects showed a reduction in BMI, serum TNF-α, and IL-6, while improving lipid profile (Bernini et al., 2016). A 2-month treatment with yogurt supplemented with a probiotic mix (Lactobacillus acidophilus La5 and Bifidobacterium lactis Bb12) in men and women with metabolic syndrome improved fasting blood glucose and insulin sensitivity. Importantly, it improved some vascular and endothelial function markers, like vascular cell adhesion molecule 1 (VCAM-1) and plasminogen activator inhibitor 1 (Rezazadeh et al., 2019). Healthy subjects with BMI at the upper limit of the healthy range randomized for a 12-week treatment with Bifidobacterium lactis with arginine supplementation appeared to have a better endothelial function and hence a reduced risk of developing atherosclerosis (Matsumoto et al., 2019). This suggests a possible therapeutic and preventive effect of probiotics on endothelial function and CV risk. However, conclusions should be drawn carefully from these results given the possible contribution of arginine supplementation to endothelial improvement and the intervention being on low-risk subjects.
Interestingly and worth investigating is the interplay between sex hormones and probiotics, as these positive outcomes seem to be independent of the status of sex hormones. In obese postmenopausal women, administration of multispecies probiotic for 12 weeks seemed to improve metabolic parameters: serum insulin, glucose, LPS, total lipid profile, uric acid, and HOMA-IR in both high and low dose groups; however, reduced adiposity was only observed in the high-dose arm, suggesting an improved gut permeability and reduced cardiometabolic risk factors (Szulińska et al., 2018). Similarly, premenopausal women diagnosed with polycystic ovarian syndrome were treated with pomegranate juice with and without probiotics mix (Lactobacillus rhamnosus GG, bacillus koagolans, and indicous) for 8 weeks. The group receiving probiotics showed an improved metabolic and inflammatory function alongside reduction in blood pressure (Esmaeilinezhad et al., 2020). Women aged 20–50 years with arterial HTN treated with a probiotic cocktail (Lactobacillus para casei LPC-37, Lactobacillus rhamnosus HN001, Lactobacillus acidophilus NCFM, and Bifidobacterium lactis HN019) for 8 weeks showed an improved fasting blood glucose, cholesterol, and elevated HDL level compared with the baseline. Interestingly, probiotics improved autonomic function and heart rate variability by reducing the low frequency domain, without significantly changing blood pressure; yet systolic BP was reduced by 5 mmHg compared with the placebo (Romão da Silva et al., 2020)
On the other hand, several clinical trials failed to record therapeutic benefits of probiotics on metabolic and cardiovascular outcomes. For instance, the commercial probiotic VSL#3,, which contains 8 different strains of lactic acid bacteria, was used in a twice daily intervention for 10 weeks in men and women with nonalcoholic fatty liver disease. It did not appear to improve cardiovascular risk factors and liver injury scores. Nevertheless, it improved HOMA-IR (Chong et al., 2021). Another randomized controlled crossover study showed that metabolic syndrome symptoms were not alleviated by a daily intervention with Lactobacillus reuteri V3401 strain for 12 weeks. Yet, this intervention was able to reduce some inflammatory markers, IL-6 and VCAM-1 (Tenorio-Jiménez et al., 2019). Although probiotics exhibited some beneficial metabolic outcomes in human trials, results seemed to be dependent on the bacterial species used. As such, some probiotics did not seem to change dysbiosis-related parameters such as gut permeability compared with the control groups (Leber et al., 2012; Ivey et al., 2015; Stadlbauer et al., 2015; Grąt et al., 2017). For instance, using Lactobacillus casei Shirota for 12 weeks in subjects with metabolic syndrome did not correct dysbiosis nor improve gut integrity (Stadlbauer et al., 2015). Thus, conclusions drawn from probiotics intervention must be specified to the strains and concentrations used. Another important note is that not only different strains have been used in probiotics studies, but even some trials used different approaches in implementing the interventions, such as using probiotics with other dietary components (Cavallini et al., 2016; Scorletti et al., 2018; Rezazadeh et al., 2019; Ruscica et al., 2019), or other dietary interventions and lifestyle modification,s such as calorie restriction and physical activity (Behrouz et al., 2017). Therefore, the role of probiotics in combating metabolic and cardiovascular insults must be carefully investigated, and studies should be accurately designed to limit other confounding factors such as dietary and lifestyle modifications. However, controlled use of probiotics can be safe and useful in preventing CVDs and metabolic derangements in low-risk individuals alone or in combination with other compounds such as prebiotics (Behrouz et al., 2017; Trotter et al., 2020).
Antibiotics
Antibiotics were proposed to be one of the interventions to achieve eubiosis (Ianiro et al., 2016). However, few studies explored the efficacy of antibiotics in ameliorating dysbiosis related cardiometabolic dysfunction. For example, one study used antibiotics to reset GM community in patients with type 2 diabetes mellitus and obesity. Yet, major metabolic parameters such as insulin sensitivity, systemic inflammation, gut permeability, and adipocyte size did not change positively in response to a 7-day treatment of amoxicillin, vancomycin, or placebo (Reijnders et al., 2016). One case study on a postmenopausal woman suffering from chronic resistant HTN for 3 years, which was uncontrolled on more than 3 antihypertensive drugs, in addition to a history of metabolic and immune pathologies including diabetes and arthritis, reported a temporary (6-month) improvement of her HTN control upon treatment with a postoperative antibiotic mix (IV vancomycin, rifampin, and ciprofloxacin orally) (Qi et al., 2015). Given the adverse effects associated with antibiotics use and the risk of development of antibacterial resistance, this might be the least desirable intervention to correct dysbiosis and related pathologies.
Fecal Microbial Transplantation
Fecal microbial transplantation (FMT) is a novel method that has been recently suggested to induce eubiosis and alleviate pathologies mediated by disturbed gut microbiome. It is the process of isolating GM from healthy donors to transplant it into diseased subjects. FMT can be done through various methods that are relatively safe and noninvasive, rectally like enema, naso-gastric route, or orally by capsules (Lagier, 2014; Wang et al., 2016). Transplanted microbiota can be homologous from the same person and heterologous/allogenic from first-degree relatives or other healthy subjects. Interestingly, allogenic GM transplantation was found to be more effective than homologous interventions (Grehan et al., 2010; Wang et al., 2016; Schepici et al., 2019). Adverse effects reported after FMT are not serious. The side effects reported were mainly abdominal discomfort and diarrhea for a few hours after the procedure (Lagier, 2014). However, there remains concerns about the potential safety/side effects related to the nonbacterial component of the fecal material transferred (Bojanova and Bordenstein, 2016). A preliminary report observed that sterile protein isolates from donor fecal material were able to induce the required response in recipients (Ott et al., 2017); however, future investigation will be required to determine the possibility of fractionation and reducing the content being transplanted to the necessary organisms only.
FMT has been considered as one of the important lines of life-saving treatments for patients with Clostridium difficile infections as these patients had improved outcomes and less chances for reoccurrence than those receiving conventional treatments (Van Nood et al., 2013; Kelly et al., 2016; Lee et al., 2016). It was even suggested that FMT can be promising in eradicating multidrug resistant microorganisms (Saha et al., 2019). Since GM has been identified as an important variable in the pathogenicity and prognosis of a large set of metabolic and cardiovascular diseases, FMT might be effective in correcting and alleviating dysbiosis related dysfunctions, especially the ones starting early on and having no clear treatment regimens.
One clinical trial on male subjects with metabolic derangements including hyperinsulinemia, BMI above 30, elevated waist circumference, and increased adiposity treated with purified GM from lean and healthy donors (allogenic transfer) matched in sex and age through a duodenal tube over a 6-week period showed improved insulin sensitivity and microbial diversity favoring butyrate producing bacteria such as Roseburia intestinalis compared with the control group receiving an autologous transfer (Vrieze et al., 2012). However, another double-blinded randomized clinical trial using FMT delivered through capsules once per week from healthy donors to subjects with obesity and insulin resistance did not record any difference in either parameter after 12 weeks of intervention (Reijnders et al., 2016). Similar results were recorded from a randomized trial of obese adolescents, after ingestion of 28 capsules of lean donors’ GM. Up to 26 weeks postintervention, recipients of either sex did not show any evidence of improvement neither in metabolic parameters such as insulin sensitivity nor in obesity, although the central to peripheral fat ratio was reduced in the FMT arm only (Leong et al., 2020). Furthermore, FMT in patients with nonalcoholic fatty liver disease who suffered from insulin resistance delivered directly to the colon from autologous and allogenic sources did not improve insulin resistance; however, allogenic FMT improved gut permeability (Craven et al., 2020). Also, FMT in patients with metabolic syndrome from a healthy vegan donor did not alleviate TMAO levels and vascular inflammation (Smits et al., 2018). Although FMT has been revolutionary in treating diseases like Clostridium difficile infections, ulcerative colitis, and others, its role in cardiometabolic dysfunction is not fully understood, requires further investigation, and its long-term efficacy has yet to be established (Zhang et al., 2019b).
Short-Chain Fatty Acids
In human studies, SCFAs were measured as a secondary outcome of dietary intervention rather than being the treatment per se, and most evidence is drawn from animal studies. One recent study using rectal capsule delivery of SCFAs in a triple-blinded randomized trial examined the effect of 1 week of SCFAs administration on psychosocial stress of 66 healthy men. In the two intervention arms, low and high SCFAs were equally successful in reducing cortisol levels in response to psychosocial stress, and both had an increased serum SCFAs as well compared with the placebo arm (Dalile et al., 2020). The OmniHeart study, which included 164 adults, assessed the role of macronutrients on serum SCFAs levels. Three isocaloric and high-fiber diets rich in either carbohydrate, protein, or unsaturated fat were applied for 6 weeks. The results indicated differences in SCFAs serum levels in response to different diets, which were correlated with some cardiometabolic aspects. For instance, the butyrate level was only increased by a high-protein diet and was associated with decreased HDL levels and ghrelin and increased insulin and glucose levels (Mueller et al., 2020). A lot of questions and concerns arise regarding the use of SCFAs for therapeutic purposes in humans, especially in cardio and metabolic pathologies. Specifically, therapeutic dose selection, safety and efficacy of single or combined use, and most importantly the long-term effects of their use. Henceforth, more clinical studies should be done using SCFAs as a therapy for early metabolic derangements in a sex-dependent fashion.
Conclusion
Dysbiosis is a common occurrence in patients suffering from cardiometabolic conditions. Not only do GM alterations in these patients appear to be driven by the same risk factors of the other pathologies, but they also seem to contribute to and drive the molecular changes leading to cardiovascular involvement, including AT inflammation, particularly in PVAT. Sexual dimorphism is evident is several steps starting at the differential effect of sex hormones on GM diversity and stability, encompassing sex-dependent effects on GM metabolite production, gut permeability, vulnerability of AT to inflammatory changes, and culminating in a different susceptibility to CVD incidence. Future investigation utilizing systematic approaches is required for a better understanding of the pathways involved to allow for tailored therapy for effective management of early cardiometabolic dysfunction in either sex.
Authorship Contributions
Wrote or contributed to the writing of the manuscript: Dwaib, AlZaim, Ajouz, Eid, El-Yazbi.
Footnotes
- Received June 14, 2021.
- Accepted October 23, 2021.
This work was supported by an American University of Beirut (AUB) Faculty of Medicine MPP grant to A.E.-Y. H.S.D. is supported by a Ph.D. Scholarship from the Faculty of Agriculture and Food Sciences at AUB and a l’Oreal–UNESCO for Women in Science Fellowship. I.A. is supported by a MasterCard Foundation Scholarship.
No author has an actual or perceived conflict of interest with the contents of this article.
Abbreviations
- Akt
- protein kinase B
- AMPK
- AMP-activated protein kinase
- AT
- adipose tissue
- AT2-R
- angiotensin 2 receptor
- BMI
- body mass index
- BP
- blood pressure
- CVD
- cardiovascular disease
- E2
- 17β-estradiol
- ERβ
- estrogen receptor-β
- F/B
- Firmicutes/Bacteroidetes
- FMO3
- flavinmonooxygenase 3
- FMT
- fecal microbial transplantation
- GM
- gut microbiota
- GPR
- G-protein coupled receptor
- HDL
- high-density lipoprotein
- HF
- heart failure
- HFD
- high-fat diet
- HOMA-IR
- homeostatic model assessment–insulin resistance
- HTN
- hypertension
- IL
- interleukin
- LPS
- lipopolysaccharide
- MUC2
- mucin 2
- MyD88
- myeloid differentiation primary response 88
- NF-κB
- nuclear factor kappa-light-chain-enhancer of activated B cells
- NOX
- NADPH oxidase
- PGC-1α
- PPARγ gamma coactivator-1α
- PLC
- phospholipase C
- PVAT
- perivascular adipose tissue
- SCFA
- short chain fatty acids
- SREBP-1
- sterol response element binding protein-1
- TLR4
- toll-like receptor 4
- TMA
- trimethylamine
- TMAO
- trimethylamine-N-oxide
- TNFα
- tumor necrosis factor α
- Tregs
- regulatory T cells
- VCAM-1
- vascular cell adhesion protein 1
- WAT
- white adipose tissue
- Copyright © 2022 by The American Society for Pharmacology and Experimental Therapeutics
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
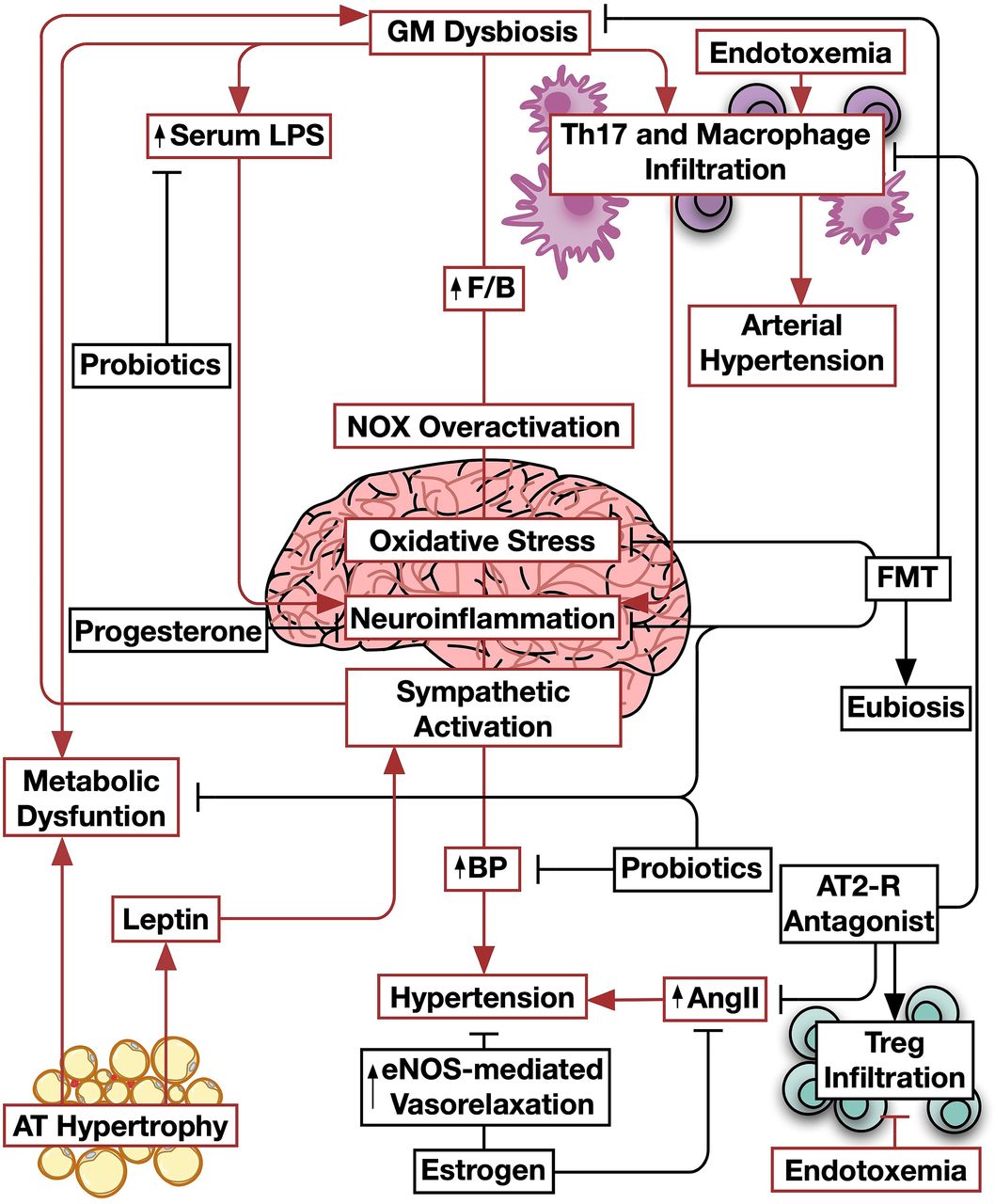{kind=link}
{kind=link}
Jump to section
- Article
- Visual Overview
- Abstract
- Introduction
- Sex-Dependent Differences in Gut Microbiota in the Healthy State
- Sex-Dependent Gut Microbiome Alterations in Early Metabolic Impairment
- Sex-Dependent Impact of Gut Microbiome on Metabolically Induced Cardiovascular Dysfunction
- Therapeutic Interventions for Cardiometabolic Consequences of Dysbiosis
- Conclusion
- Authorship Contributions
- Footnotes
- Abbreviations
- References
- Figures & Data
- Info & Metrics
- eLetters