



Abstract
We tested the hypothesis that isoform shifts in sarcomeres of the immature heart modify the effect of cardiac myosin-directed sarcomere inhibitors and activators. Omecamtiv mecarbil (OM) activates tension and is in clinical trials for the treatment of adult acute and chronic heart failure. Mavacamten (Mava) inhibits tension and is in clinical trials to relieve hypercontractility and outflow obstruction in advanced genetic hypertrophic cardiomyopathy (HCM), which is often linked to mutations in sarcomeric proteins. To address the effect of these agents in developing sarcomeres, we isolated heart fiber bundles, extracted membranes with Triton X-100, and measured tension developed over a range of Ca2+ concentrations with and without OM or Mava treatment. We made measurements in fiber bundles from hearts of adult nontransgenic (NTG) controls expressing cardiac troponin I (cTnI), and from hearts of transgenic (TG-ssTnI) mice expressing the fetal/neonatal form, slow skeletal troponin I (ssTnI). We also compared fibers from 7- and 14-day-old NTG mice expressing ssTnI and cTnI. These studies were repeated with 7- and 14-day-old transgenic mice (TG-cTnT-R92Q) expressing a mutant form of cardiac troponin T (cTnT) linked to HCM. OM increased Ca2+-sensitivity and decreased cooperative activation in both ssTnI- and cTnI-regulated myofilaments with a similar effect: reducing submaximal tension in immature and mature myofilaments. Although Mava decreased tension similarly in cTnI- and ssTnI-regulated myofilaments controlled either by cTnT or cTnT-R92Q, its effect involved a depressed Ca2+-sensitivity in the mature cTnT-R92 myofilaments. Our data demonstrate an influence of myosin and thin-filament associated proteins on the actions of myosin-directed agents such as OM and Mava.
SIGNIFICANCE STATEMENT The effects of myosin-targeted activators and inhibitors on Ca2+-activated tension in developing cardiac sarcomeres presented here provide novel, ex vivo evidence as to their actions in early-stage cardiac disorders. These studies advance understanding of the molecular mechanisms of these agents, which are important in preclinical studies employing sarcomere Ca2+-response as a screening approach. The data also inform the use of commonly immature cardiac myocytes generated from human-inducible pluripotent stem cells in screening for sarcomere activators and inhibitors.
Introduction
A recent advance in the treatment of adult cardiac disorders is the development of inotropic agents that are new classes of sarcomere activators and inhibitors in phase 3 clinical trials. The successful and safe use of these agents in adult hearts led to our hypothesis that they may also be effective in immature myofilaments of pediatric patients with heart failure and cardiomyopathies. The agents we employed in our studies of Ca2+-activation of myofilament tension are omecamtiv mecarbil (OM) [C20H24FN5O3, methyl 4-(2-fluoro-3-(3-(6-Methylpyridin-3-yl)ureido)benzyl)piperazine-1-carboxylate], a myosin-targeted sarcomere activator (Teerlink et al., 2020; Teerlink et al., 2021) (Fig. 1A), and mavacamten (Mava) [C15H19N3O2, (S)-3-Isopropyl-6-((1-phenylethyl)amino) pyrimidine-2,4(1H,3H)-dione], a myosin-targeted sarcomere inhibitor (Toepfer et al., 2020; Zampieri et al., 2021) (Fig. 1B). OM was developed to enhance sarcomere response to Ca2+ without affecting myocyte Ca2+ fluxes (Malik et al., 2011; Planelles-Herrero et al., 2017). OM is currently in clinical trials testing its effects in heart failure patients with reduced ejection fraction (Teerlink et al., 2021). Mava was developed as a sarcomere inhibitor for use in hypertrophic cardiomyopathy (HCM) with hypercontractility and outflow obstruction, which is often linked to sarcomere protein mutations. Mava is also in a clinical trial testing its effects in patients with HCM, a condition that may lead to heart failure with preserved ejection fraction (Ho et al., 2020).

Chemical structure depiction of (A) omecamtive mecarbil (source: https://pubchem.ncbi.nlm.nih.gov/compound/Omecamtiv-mecarbil) and (B) mavacamten (source: https://pubchem.ncbi.nlm.nih.gov/compound/Mavacamten).
Cardiac contraction and relaxation are controlled by sarcomeres expressing different protein isoforms in mature and immature myofilaments. A dominant isoform switch occurs in cardiac troponin, a thin filament heterotrimeric complex that, together with tropomyosin (Tm), controls the Ca2+-dependent reaction of thin filaments with force-generating myosin cross-bridges (Solaro et al., 2013; Solis and Solaro, 2021). The mechanism of action for OM and Mava has been attributed to their modification of the state of myosin cross-bridges, effectively increasing or decreasing the number of myosin heads that can participate in shortening and force generation (Green et al., 2016; Teerlink et al., 2021). However, evidence exists that the mechanism may also involve other sarcomere proteins (Mamidi et al., 2015; Kampourakis et al., 2018), including those that shift isoforms in the immature heart. A significant isoform shift occurs in the thin filament regulatory unit made up of 7 actins, with one Tm and one troponin complex consisting of cardiac troponin C (cTnC) (a Ca2+-binding unit), troponin I (TnI) (an inhibitory unit), and TnT (a Tm-binding unit). Ca2+-binding to cTnC triggers contraction by releasing the actin–cross-bridge interaction from the combined inhibition of cardiac TnI (cTnI), cTnT, and Tm. Promotion of actin–cross-bridge reactions also occurs in a highly cooperative process involving near-neighbor interactions of regulatory units. Although there are similar control mechanisms in immature and mature hearts, adult sarcomeres are controlled by an adult form, cTnI, whereas immature sarcomeres are controlled by a fetal/neonatal isoform, which is identical to the slow skeletal troponin I (ssTnI) isoform (Hunkeler et al., 1991; Murphy et al., 1991). The ssTnI isoform is expressed in fetal and neonatal hearts before a complete switch to the adult cTnI. In mouse hearts, the switch from ssTnI occurs incrementally with the expression of 100% cTnI by 10–14 days of age. Studies of expression of ssTnI in human hearts have demonstrated a similar pattern in developing and adult human hearts (Sasse et al., 1993).
Among the changes in the isoform population that occur in the fetal/neonatal heart, there is substantial evidence that the impact of ssTnI on the control of cardiac myofilaments is a dominant mechanism in developing hearts (Metzger et al., 2003; Layland et al., 2005). In addition to isoform distributions, molecular modifications in the primary structure of ssTnI also alter pH sensitivity, cooperative activation, and protein phosphorylation (Solaro et al., 2013). Cardiac muscle preparations with sarcomeres controlled by ssTnI demonstrate altered responses to physiologic controllers (Solaro et al., 2013), pathologic stresses, and pharmacological interventions (Arteaga et al., 2000), compared to cTnI-controlled sarcomeres. These findings indicate a strong possibility that inhibition or activation of sarcomeres by OM and Mava may be different in immature compared with adult sarcomeres.
We tested the effects of OM and Mava on myofilaments from adult and neonatal hearts expressing either cTnI and ssTnI together with wild-type cTnT, or a mutant cTnT linked to HCM. Our results revealed novel differential effects of the agents on Ca2+ sensitivity and cooperative activation of tension in sarcomeres controlled by either cTnI or ssTnI. These results provide evidence relating to the effects of these sarcomere-directed agents in early-stage cardiac disorders.
Methods
Mouse Models
Experiments were approved by the Animal Care and Use Committee of the University of Illinois at Chicago and in compliance with the Guide for the Care and Use of Laboratory Animals as adopted by the U.S. National Institutes of Health. One set of experiments employed male and female adult non-transgenic (NTG) or transgenic (TG) adult mice on an FVB/N mouse, named after its susceptibility to Friend leukemia virus B (FVB/N) background expressing ssTnI (TG-ssTnI) in the cardiac compartment, as previously described (Alves et al., 2017). A second set of experiments employed male and female 7- and 14-day-old NTG mice and TG mice (TG-cTnT-R92Q) expressing an HCM-linked TnT mutant generated in C57/B6 strain as previously described (Tardiff et al., 1999; Chowdhury et al., 2020). Since it is well-known that genetic background may have a notable effect on the HCM phenotype (Prabhakar et al., 2001; Michele et al., 2002; Rowlands et al., 2017), we first rederived and characterized TnT-R92Q mice in FVB/N background to be consistent with TG-ssTnI mice. We crossed TnT-R92Q heterozygous males with FVB/N females to create F1 generation of mice. Next, we bred TnT-R92Q-positive males from F1 generation with FVB/N females to obtain F2 generation of mice. We repeated this process up to F10 generation.
Detergent-Extracted (Skinned) Bundles of Ventricular Muscle Fibers
In one set of experiments, we extracted hearts from 3–5-month-old NTG and TG-ssTnI mice that were heparinized and anesthetized with a ketamine/xylazine solution (200mg/20mg/kg body weight) in accord with the Veterinary Medical Association Panel on Euthanasia Guidelines. To prepare detergent-extracted fiber bundles, we carried out procedures essentially as previously described (Ryba et al., 2019). Following heart extraction and exposure of the left ventricular papillary muscle, we dissected fiber bundles 3–5 mm in length and 150–200 µm in diameter in a high-relaxing buffered solution (HR), pH 7.0 [10 mM EGTA, 41.89 mM potassium propionate, 100 mM N,N-bis(2-hydroxyethyl)-2-aminoethanesulfonic acid, 6.75 mM magnesium dichloride, 6.22 mM adenosine 5′-triphosphate disodium salt hydrate, 10 mM disodium creatine phosphate, 5 mM sodium azide, ionic strength 150 mM].
Use of sodium azide is standard practice in skinned fiber experiments, as in some protocols acto-myosin ATPase rate and cTnC Ca2+-binding is measured simultaneously with force. Azide prevents mitochondrial ATP hydrolysis, which would mask the sarcomere ATPase rate and inhibit mitochondrial Ca2+-binding and uptake. Together with solubilization of mitochondria with Triton X-100, sodium azide assures that the measurements reflect the activity of sarcomeres. The solutions employed in the skinned fiber experiments contain high physiologic levels of MgATP that is buffered with phosphocreatine (creatine phosphate). Creatine phosphate is important for proper buffering of the sarcomeric ATP and ADP changes that would otherwise affect contractile activity. Addition of creatine phosphokinase is required to catalyze the phosphate transfer from creatine phosphate to ADP and provides a supplement to the endogenous creatine kinase (Roche et al., 2015).
Protease inhibitors [pepstatin A 2.5 µg/ml; leupeptin, 1 µg/ml; phenylmethylsulphonyl fluoride (PMSF), 50 µmol/l] were included in solutions. Following isolation, fiber bundles were immersed in 1% Triton X-100 in HR for 30 minutes at room temperature and then returned to HR without Triton. After the detergent skinning, fibers were transferred to HR solution and kept briefly on ice. Fibers were mounted between the arm of a micromanipulator and a force transducer and immersed in HR solution containing 0.06% dimethylsulfoxide (DMSO, the drug vehicle). Sarcomere length was adjusted with a micromanipulator to 2.2 µm using the diffraction pattern generated from a helium-neon laser and kept constant during the protocol. In a second set of experiments, we carried out the same procedures with fiber bundles from 7- and 14-day-old NTG mice and TG mice expressing cTnT-R92Q. In this case the 7-day-old mice were decapitated, and the 14-day-old mice were anesthetized with isoflurane before extracting the hearts. Fiber bundles were then skinned in 1% Triton X-100 for either 10 minutes (7-day) or 20 minutes (14-day).
Determination of Ca2+ Dependence of Tension Developed in Skinned Fibers
Tension developed in the skinned fibers was measured over a range of free Ca2+ concentrations expressed as pCa (−log [Ca2+]) values. The pCa solutions were prepared by mixing various ratios of HR with 9.98 mM CaCl2 (pCa 4.5). We added 0.1 µM creatine phosphokinase in all solutions; addition of creatine phosphokinase supplements the endogenous kinase to ensure transfer of phosphoryl group from creatine phosphate to ADP (Roche et al., 2015). The solutions also contained 0.06% DMSO, 0.3 µM OM, or 0.3 µM Mava in 0.06% DMSO. Ratios of HR and 9.98 mM CaCl2 were used to generate a range of pCa 7.0–5.5 were calculated using WEBMAXC Standard. All experiments were performed at 22°C, as chemically skinned fibers are more stable in subphysiologic temperature, which further allows reproducibility of the measurements (Lamb and Stephenson, 2018). We initially induced maximum tension at pCa 4.5, relaxed the fibers in HR with DMSO, and determined the diameter and width of the fiber bundle at 3 points for calculation of cross-sectional area. After a 5-minute preincubation period in HR-DMSO, we recorded developed tension over the range of pCa values with a chart recorder. Following a 5-minute preincubation with HR containing either 0.3 µM OM or 0.3 µM Mava, the measurements were repeated with the same fiber bundle in the range of pCa solutions containing either Mava (0.3 µM) or OM (0.3 µM). These drug concentrations are based on our previous studies, as well as reports from other laboratories investigating force development in skinned fibers from human and animal hearts (Malik et al., 2011; Utter et al., 2015; Green et al., 2016; Planelles-Herrero et al., 2017; Mamidi et al., 2018; Sparrow et al., 2020; Awinda et al., 2021). Skinned fibers were checked for maintenance of maximum tension. The pCa tension data were fit to the following equation:
 pCa50 is the negative logarithm of [Ca2+] at half-maximum tension generation, nH is the Hill coefficient, Y0 is the baseline, and A is the amplitude.
pCa50 is the negative logarithm of [Ca2+] at half-maximum tension generation, nH is the Hill coefficient, Y0 is the baseline, and A is the amplitude.
Gel Electrophoresis and Western Blot Analysis
To separate troponin I isoforms, 12% sodium dodecyl phosphate polyacrylamide gel electrophoresis (SDS-PAGE) was performed and transferred to an Immun-Blot PVDF membrane (BioRad) as previously described (Warren et al., 2010) with BioRad’s Criterion gel box and blotter. To separate troponin T isoforms and the R92Q mutant 8% total acrylamide, 1.1% Bis-acrylamide cross-linked SDS-PAGE gels (Anderson et al., 1991) were run in a large-format gel box (SE600 Hoefer). The large gel was cut down to fit into a BioRad Criterion blotter to transfer the protein onto Immun-Blot PVDF (BioRad) to perform western blot analysis. The whole homogenate samples for this study were prepared as described (Batra et al., 2021). The troponin separations were loaded at 10 µg/lane and the myosin heavy chain (MHC) separations were loaded at 2.7 µg/lane. The protein concentrations for all samples were determined with Thermo Scientific’s 660nm protein assay with the ionic detergent compatibility reagent added. The myosin heavy chain isoform separation was performed as previously described (Warren and Greaser, 2003) and the bands were detected with Coomassie G-250. Image acquisition for both Western blots and Coomassie stained gels was done with BioRad’s Chemidoc MP imager. Gel bands and Western blot densitometric analysis was done with Image laboratory v6.01 (BioRad) and ratio analysis was done with Microsoft Excel 365.
Data and Statistical Analysis
The experiments performed in this study were exploratory in nature and did not employ a predefined plan, thus P values are descriptive. Tension was calculated as force per cross-sectional area. We fit the relation between developed isometric tension pCa by a nonlinear least squares regression analysis to the Hill’s equation as described above. A two-way analysis of variance with a Tukey post hoc adjusted P value for multiple comparisons method was used (GraphPad Prism v9.1.2) to compare maximum tension, pCa50, Hill slope, and protein isoform distributions as indicated in figure and table legends. Data exclusion criteria were set in advance utilizing the ROUT outlier test in GraphPad Prism v9.1.2 with the Q-value set at 0.05. Data sets for two-way ANOVAs were normally distributed as assessed in GraphPad Prism v9.1.2 by Shapiro-Wilk or D’Agostino-Pearson omnibus (K2). In addition, homogeneity of variance was tested by Brown-Forsythe and Bartlett’s tests. Where binary comparisons were appropriate, an unpaired two-tailed t test was performed where all data were normally distributed, but if the normality and equal variances assumptions were violated, we performed a Mann-Whitney test or an unpaired two-tailed t test with Welch’s correction respectively. For the comparison of changes in tension expressed as a percent of control tension, a pairwise comparison between untreated and treated fibers was performed, followed by an unpaired two-tailed t test between only the treated fibers. Data are presented as means ± standard deviation with a level of significance set at P < 0.05.
Results
Protein Isoform Profiles of Skinned Fiber Bundles
To investigate the effect of the presence of the fetal/neonatal isoform ssTnI on pCa-tension relations of skinned fibers treated either with vehicle or with a sarcomere activator (OM) and a sarcomere inhibitor (Mava), we employed skinned fibers from hearts of adult NTG mice and adult TG mice harboring a transgene expressing ssTnI in place of cTnI in the cardiac compartment in the first set of experiments. Previous studies have documented that the expression and stoichiometry of the sarcomeric proteins, including TnI, are unaffected in the TG-ssTnI sarcomeres compared with controls (Fentzke et al., 1999). We also employed fibers from hearts of 7- and 14-day-old NTG or TG-cTnT-R92Q mice naturally expressing variations in ssTnI during development with and without expression of an HCM-linked mutant cTnT. Figure 2 shows western blot analysis probing for cTnI, ssTnI, cTnT, and cTnT-R92Q in whole heart homogenates. As illustrated in Fig. 2, A and B, the adult TG-ssTnI hearts expressed 100% ssTnI, whereas NTG littermates expressed 100% cTnI. In 7-day-old NTG hearts, percent abundance of ssTnI was 55.3%, and the 7-day-old TG-cTnT-R92Q hearts were 63.4% (Fig. 2, A–C; Supplemental Fig. 1). There was no detectable ssTnI isoform in 14-day-old NTG or TG-cTnT-R92Q mice. The abundance of cTnT-R92Q in cTnT-R92Q hearts at 14 days was 74.7% versus 47.1% in the 7-day-old cTnT-R92Q hearts (Fig. 2, D–F; Supplemental Fig. 1). The myosin heavy chain (MHC) isoform population, detected by SDS-PAGE Coomassie stained gels, was unaffected by the presence of ssTnI or cTnT-R92Q mutant (Fig. 2, G–I). As expected, we observed statistically significant changes in the population of MHC isoforms (α and β) with aging from 7 to 14 days (Fig. 2, G and I).

Western blot and SDS-PAGE analysis demonstrating myofilament isoform abundance differences in adult and immature NTG and TG-cTnT-R92Q mouse hearts. (A) Western blot probing for troponin I isoforms in adult samples, cardiac troponin I, and slow skeletal troponin I, with an antibody recognizing both isoforms simultaneously. (B) Histograms quantifying the percent of TG-ssTnI in the total TnI population of adult samples from panel (A). (C) Histograms quantifying the percent of TG-ssTnI in the total TnI population of 7- and 14-day-old samples from panel (A). (D) Western blot probing for TnT isoforms with an antibody recognizing all isoforms simultaneously. (E) Histograms quantifying the percent of TG-cTnT-R92Q in the total TnT population of adult samples from panel (D). (F) Histograms quantifying the percent of TG-cTnT-R92Q in the total TnT population of 7- and 14-day-old samples from panel (D). (G) Representative SDS-PAGE stained with Coomassie blue showing the myosin heavy chain (MHC) α and β isoforms. (H) Histograms quantifying the percent of β-MHC in the total MHC in adult samples from panel (G). Statistical significance was determined by an unpaired, two-tailed T-test. No significance was found. (I) Histograms quantifying the percent of β-MHC in the total MHC in 7- and 14-days old samples from panel (G). Data presented as mean ± S.D., n = 4, **P < 0.0042, ****P < 0.0001. Statistical significance was determined by two-way ANOVA test, followed by a Tukey’s multiple comparison post hoc test for panels C, F, and I. No statistical test was performed for panels (B) or (E). Day, age of animals in days; WT, wild-type.
Studies with Omecamtiv Mecarbil
Figure 3A shows pCa-tension relations of skinned fiber bundles from hearts of 4-month-old adult control NTG and TG-ssTnI mice with and without treatment with OM. In these experiments, we treated the skinned fibers with 0.3 µM OM based on dose response relations determined in the present experiments and in our previous publications (Utter et al., 2015; Broughton et al., 2016). The parameters describing the pCa-tension relations are maximum tension (Fig. 3B; Table 1), Ca2+-sensitivity as quantified by the pCa50 (−log of the half-maximally activating molar Ca2+ concentration) (Fig. 3C; Table 1), and the steepness of the curves, reflecting cooperative activation (Arteaga et al., 2000; Solis and Solaro, 2021) and quantified by nH (Fig. 3D; Table 1). There was a difference in maximum tension developed by the skinned fibers from NTG and TG-ssTnI hearts: NTG developed 25.85% higher maximal value (P = 0.0319), but OM had no effect on these tensions (Fig. 3B; Table 1). Without OM, as expected (Fentzke et al., 1999; Arteaga et al., 2000), myofilaments controlled by ssTnI were more sensitive (by 3.74%) to Ca2+ compared with myofilaments controlled by cTnI (P = 0.0002) (Fig. 3C; Table 1). OM increased the pCa50 in both NTG, by 2.81% (P = 0.035), and TG-ssTnI fibers, by 2.20% (P = 0.0164). Although the change (Δ) in pCa50 induced by OM was 0.16 pCa units in the NTG and 0.13 in the TG-ssTnI myofilaments, (Fig. 3C; Table 1), the Ca2+ sensitivity of the OM-treated TG-ssTnI fibers was 3.12% greater than the Ca2+ sensitivity of the OM-treated NTG fibers (P = 0.0164 versus P = 0.035). Without OM, compared with the NTG controls, TG-ssTnI fibers demonstrated a 44.75%-smaller slope of the Ca2+-tension relation (P < 0.0001) (Fig. 3D; Table 1). OM induced a 2.72-fold greater depression (Δ nH = 3.16) in NTG compared with TG-ssTnI fibers (Δ nH = 1.16), such that the nH in the presence of OM was 21.54% lower in NTG when compared with TG-ssTnI fibers (P = 0.0027) (Fig. 3D; Table 1). To determine more clearly the effect of OM on the skinned fibers, we plotted the percent of control tension over a range of pCa values likely to be achieved physiologically (Fig. 3E; Supplemental Table 1). This analysis revealed that the percent increases in tension at these pCa values were higher in the NTG compared to TG-ssTnI fibers treated with OM by 427.40% at pCa 6.00 (P = 0.0082), 137.40% at pCa 5.85 (P = 0.0266), and by 48.3% at pCa 5.75 (P = 0.0079).

Effect of OM on pCa-tension relations of skinned fiber bundles from hearts of adult control NTG mice and TG-ssTnI mice. (A) pCa-tension relations in NTG and TG adult mouse skinned fiber bundles ± OM treatment. (B) Scatter plot of maximum tension (mN/mm2). (C) Scatter plot of pCa50 values. (D) Scatter plot of Hill slopes (nH). (E) Activation represented as OM-induced percent increase in control tension over a range of submaximal pCa values. Data presented as mean ± S.D., n = 5, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Exact P values are found in Table 1. Statistical significance was determined by two-way ANOVA test, followed by a Tukey’s multiple comparison post hoc test (A–D); and by unpaired two-tailed Student’s t test for (E).
Ca2+-activated tension parameters for skinned fibers of adult TG-ssTnI mice and NTG controls treated with 0.3 µM OM
Data are presented as mean ± S.D. Two-way ANOVA test followed by a Tukey’s multiple comparison post hoc test.
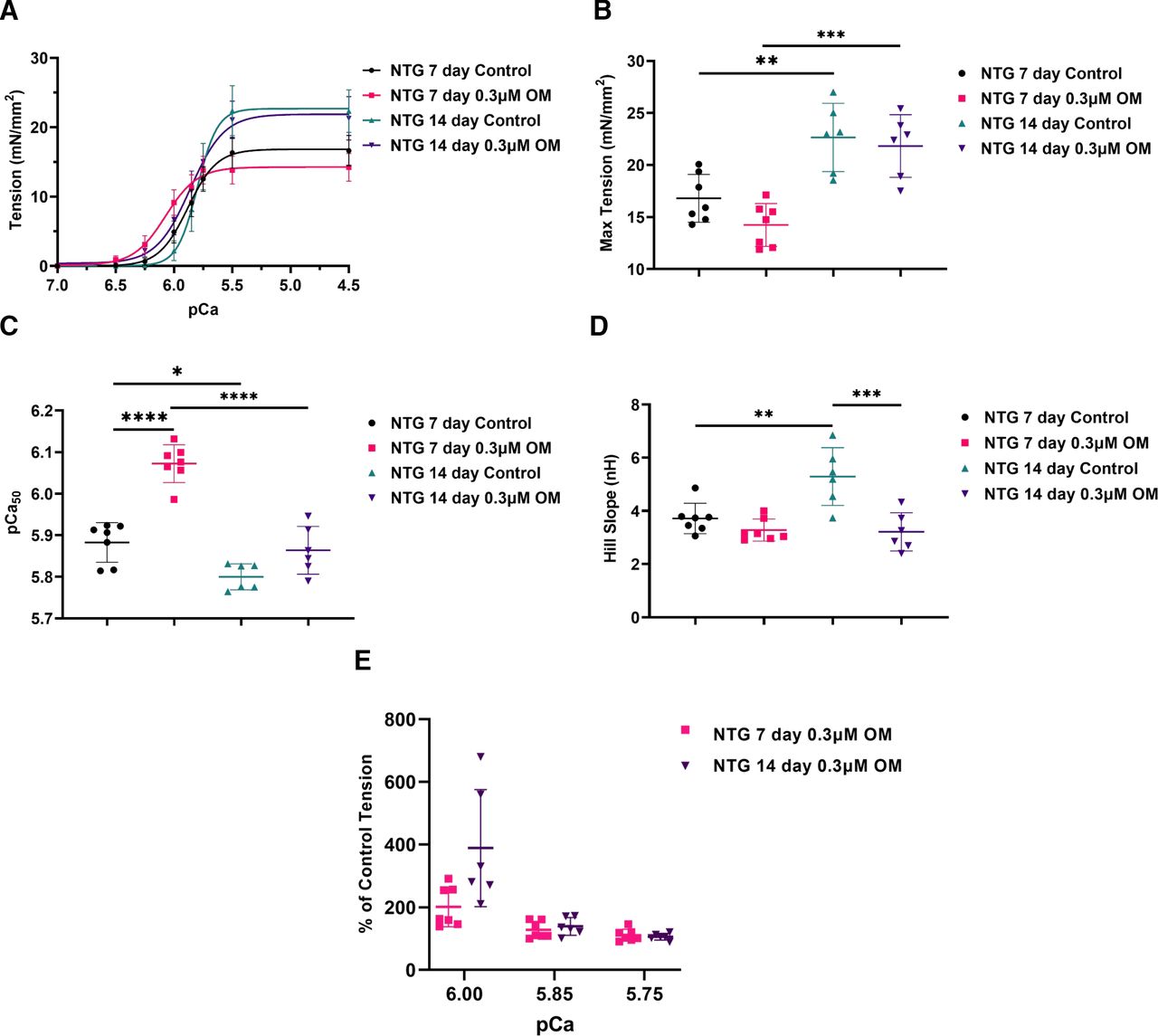
To test the effect of OM on fiber bundles from neonatal mouse hearts demonstrating natural switching from ssTnI to cTnI as illustrated in Fig. 2, we performed experiments on skinned fibers from 7- and 14-day-old mouse hearts. Figure 4A compares the pCa-tension relations of these fibers with and without additions of 0.3 µM OM. Without OM, the maximum tension developed by the 7-day fibers was 25.83% lower than the maximum tension developed by the 14-day fibers (P = 0.0035); treatment with OM had no effect on these tensions (Fig. 4B; Table 2). Without OM, the Ca2+ sensitivity of 7-day fibers was 1.43% greater than the Ca2+ sensitivity of the 14-day fibers (P = 0.0202) (Fig. 4C; Table 2). However, whereas there was a clear effect of 0.3 µM OM on pCa50 of fibers from 7-day-old hearts (P < 0.0001) by 3.21%, there was a nonsignificant effect (increase by 1.10%) of OM on pCa50 of fibers from 14-day-old hearts (Fig. 4C; Table 2). Without OM, the nH values were 29.77% lower in the 7-day-old fibers (P = 0.0037) compared with the 14-day-old fibers (Fig. 4D; Table 2). Treatment with OM did not induce a change in nH values in the 7-day-old fibers, but OM did induce a 39.30% decrease in the nH values in 14-day-old skinned fibers (P = 0.0003) (Fig. 4D; Table 2). Analysis of percent changes in tension induced by OM over a range of physiologic pCa values demonstrated a trend toward a greater relative effect of OM at pCa 6.0 that did not reach statistical significance between treated groups. There were similar percent changes in tension at other pCa values (Fig. 4E; Supplemental Table 1). In summary, in 7-day-old immature skinned fibers expressing different myofilament isoforms than the 14-day-old fibers, there were different effects of OM on Ca2+ sensitivity and cooperative activation. Yet OM induced similar relative increases in tension over the physiologic range of pCa values.

Effect of OM on pCa-tension relations of skinned fiber bundles from hearts of 7- and 14-day-old NTG mice. (A) pCa-tension relations of NTG 7- and 14-day-old mouse skinned fiber bundles ± OM treatment. (B) Scatter plot of maximum tension (mN/mm2). (C) Scatter plot of pCa50 values. (D) Scatter plot of Hill slopes (nH). (E) Activation represented as percent increase in control tension over a range of submaximal pCa values. Data presented as mean ± S.D., n = 6–7, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Exact P values are found in Table 2. Statistical significance was determined by two-way ANOVA test followed by a Tukey’s multiple comparison post hoc test (A–D); and by unpaired two-tailed Student’s t test for (E).
Ca2+-activated tension parameters for skinned fiber bundles of 7- and 14-day-old NTG mice treated with 0.3 µM OM
Data are presented as mean ± S.D. Two-way ANOVA test followed by a Tukey’s multiple comparison post hoc test.
Studies with Mavacamten
In the next series of experiments, illustrated in Fig. 5 and summarized in Table 3 and Supplemental Table 1, we compared the effects of the sarcomere inhibitor, Mava, on pCa-tension relations in skinned fibers isolated from adult hearts of NTG controls and TG-ssTnI mice. Figure 5A illustrates the results of experiments testing the effect of Mava on pCa-tension relations of the NTG and TG-ssTnI myofilaments. The results demonstrate the sarcomere-inhibiting activity of Mava by the fall in maximum tension in skinned fibers, which was nearly the same in myofilaments regulated by cTnI (47% inhibition, P = 0.0001) as those regulated by ssTnI (52% inhibition, P < 0.0001) (Fig. 5, B and E; Table 3). Mava decreased the Ca2+ sensitivity of the TG-ssTnI fibers somewhat more (decrease by 2.55%) with a ΔpCa50 of 0.15 units, whereas in NTG fibers (decrease by 2.17%) the ΔpCa50 was 0.12 units, which did not reach significance (Fig. 5C; Table 3). Mava did not induce changes in the nH values of either the TG-ssTnI or NTG fibers compared to the controls (Fig. 5D; Table 3). Data shown in Fig. 5E and summarized in Supplemental Table 1, show that the inhibition of tension shown as percent change was similar across the sub-maximum pCa values in the physiologic range.

Effect of Mava on pCa-tension relations of skinned fiber bundles from hearts of adult control NTG mice and TG-ssTnI mice. (A) pCa-tension relations in NTG and TG-ssTnI adult skinned fiber bundles ± Mava treatment. (B) Scatter plot of maximum tension (mN/mm2). (C) Scatter plot of pCa50 values. (D) Scatter plot of Hill slopes (nH). (E) Inhibition represented as percent of control tension over a range of submaximal pCa values. Data represented as mean ± S.D., n = 5–6, *P < 0.05, ***P < 0.001, ****P < 0.0001. Exact P values are found in Table 3. Statistical significance was determined by two-way ANOVA test followed by a Tukey’s multiple comparison post hoc test (A–D); and by unpaired two-tailed Student’s t test for (E).
Ca2+-activated tension parameters for skinned fiber bundles of adult TG-ssTnI mice and NTG controls treated with 0.3 µM Mava
Data are presented as mean ± S.D. Two-way ANOVA test followed by a Tukey’s multiple comparison post hoc test.
To further probe and compare the effects of Mava on sarcomeres controlled by cTnI and ssTnI, we tested its effects on skinned fibers isolated from perinatal hearts of 7- and 14-day-old NTG mice and TG mice expressing cTnT-R92Q, a mutant linked to HCM (Figs. 6 and 7; Table 4–5). Data in Fig. 6, summarized in Table 4, compare pCa-tension relations in skinned fibers from NTG controls and TG-cTnT-R92Q 7-day-old hearts with and without treatment with Mava. Without Mava, there were no differences in maximum tension, pCa50, or nH values between the 7-day-old NTG and 7-day-old TG-cTnT-R92Q fibers (Fig. 5D and 6A; Table 4). Treatment with Mava reduced the maximum tension in both groups (by 50.65% for NTG, P = 0.0186; and by 47.62% for TG-cTnT-R92Q, P = 0.0004) (Fig. 6, A and B) with no effect on the pCa50 (Fig. 6C; Table 4) or nH values (Fig. 6D; Table 4). The depression in tension induced by Mava was ∼50% over a range of submaximal pCa values (Fig. 6E; Supplemental Table 1). Data in Fig. 7, summarized in Table 5 and Supplemental Table 1, compare pCa-tension relations in skinned fibers from NTG controls and TG-cTnT-R92Q 14-day-old hearts with and without treatment with Mava. Without Mava, there was no difference between maximum tension developed by NTG and TG-cTnT-R92Q 14-day-old fibers (Fig. 7, A–B; Table 5). However, in contrast to the 7-day-old data, pCa50 was 2.88% higher in the TG-cTnT-R92Q fibers than in the NTG fibers at 14 days without Mava (P = 0.0022) (Fig. 7C; Table 5). At 14 days of age, TG-cTnT-R92Q fibers also had 46.34% lower nH values compared with the NTG fibers (P < 0.0001) (Fig. 7D; Table 5). Treatment with Mava reduced the maximum tension similarly in the 14-da-old NTG (reduction by 49.68%, P = 0.0013) and in TG-cTnT-R92Q (reduction by 43.96%, P = 0.0025) (Fig. 7, A and B; Table 5). Whereas there was no effect of Mava on the pCa50 of the 14-day-old NTG fibers (Fig. 7C; Table 5), Mava induced a 3.16% decrease in the pCa50 of the TG-cTnT-R92Q fibers (P = 0.0034) (Fig. 7C; Table 5). Values of nH were not changed by Mava in either the 14-day-old NTG or TG-cTnT-R92Q fibers (Fig. 7D; Table 5). The depression in tension at submaximal pCa values induced by Mava ranged from 11.09% in the 14-day-old NTG fibers to 38.15% in the 14-day-old TG-TnT-92Q fibers (at pCa 5.85 P = 0.0251; at pCa 5.75 P = 0.0210) (Fig. 7E; Supplemental Table 1). In summary, these data indicate that at 7 days, Mava sarcomere inhibition occurs via a depression in tension generation with no effect on pCa50 or nH values, and independently of the presence of ssTnI and cTnT-R92Q in the myofilaments. However, at 14 days, when ssTnI has been replaced by cTnI, and when a larger proportion of cTnT has been replaced by cTnT-R92Q than at 7 days, Mava induced a greater relative depression in NTG versus TG fiber tension at submaximal pCa values in conjunction with an effect to reduce the pCa50 only in the cTnT-R92Q fiber bundles.

Effect of Mava on pCa-tension relations of skinned fiber bundles from hearts of 7-day-old NTG and 7-day-old TG-cTnT-R92Q mice. (A) pCa-tension relations of NTG 7-day-old mouse skinned fiber bundles ± Mava treatment. (B) Scatter plot of maximum tension (mN/mm2). (C) Scatter plot of pCa50 values. (D) Scatter plot of Hill slopes (nH). (E) Inhibition represented as percent of control tension over a range of submaximal pCa values. Data presented as mean ± S.D., n = 3–7, *P < 0.05, ***P < 0.001. Exact P values are found in Table 4. Statistical significance was determined by two-way ANOVA test followed with a Tukey’s multiple comparison post hoc test (A, C–D); and by unpaired two-tailed Student’s t test for (E).

Effect of Mava on pCa-tension relations of skinned fiber bundles from hearts of 14-day-old NTG mice and 14-day-old TG-cTnT-R92Q mice. (A) pCa-tension relations of NTG 14-day-old skinned fiber bundles ± Mava treatment. (B) Scatter plot of max tension (mN/mm2). (C) Scatter plot of pCa50 values. (D) Scatter plot of Hill slope (nH). (E) Inhibition represented as percent of control tension over a range of submaximal pCa values. Data represented as mean ± S.D., n = 4–5, *P < 0.05, **P < 0.01, ****P < 0.0001. Exact P values are found in Table 5. Statistical significance was determined by two-way ANOVA test followed by a Tukey’s multiple comparison post-hoc test (A–D); and by unpaired two-tailed Student’s t test for (E). Data presented as mean ± S.D.
Ca2+-activated tension parameters for skinned fiber bundles of 7-day-old TG-cTnT-R92Q mice and NTG controls treated with 0.3 µM Mava
Data are presented as mean ± S.D. Two-way ANOVA test followed with a Tukey’s multiple comparison post hoc test.
Ca2+-activated tension parameters for skinned fiber bundles of 14-day-old TG-cTnT-R92Q mice and NTG controls treated with 0.3 µM Mava
Data are presented as mean ± S.D. Two-way ANOVA test followed with a Tukey’s multiple comparison post hoc test.
Discussion
Mechanisms of OM Activation and Mava Inhibition Involve Integrated Effects of Sarcomere Proteins Beyond Myosin
Our studies of the effects of OM and Mava on Ca2+-activation of skinned ventricular fiber bundles expressing either cTnI or ssTnI provide novel insights into their mechanisms of action. These mechanisms are relevant to the development of new sarcomere inhibitors and activators likely to be screened to improve therapeutic index and pharmacokinetic properties. Abundant evidence demonstrates that a key step in screening sarcomere activators and inhibitors is an in vitro determination of sarcomere response to Ca2+. The effect of Mava and OM to modify Ca2+-force relations of skinned fiber preparations encouraged development of the next stage, eventually leading to studies in myocytes, hearts, animal models, and human safety trials (Morgan et al., 2010; Green et al., 2016). Our data are also relevant to the use of immature cardiac myocytes, often expressing ssTnI, derived from human-inducible pluripotent stem cells and proposed for use in drug development and diagnosis (Wheelwright et al., 2020; Andrysiak et al., 2021).
To illuminate mechanisms affected by OM and Mava, Fig. 8 depicts thin and thick filament control of myofilament force generation. Included in Fig. 8 are recent revelations on the dispositions of the cross-bridges in two-headed myosin in a super relaxed state (SRX) with the heads folded back and in a disordered relaxed state (DRX), poised for interaction with the thin filaments (Anderson et al., 2018; Toepfer et al., 2020). In addition, an important revelation is the evidence that both TnI and TnT are in an extended state interacting with all the actins in a regulatory unit, and thus likely to be involved not only in the Ca2+ switch but also in the near-neighbor interactions among the regulatory units (Yamada et al., 2020; Solis and Solaro, 2021; Tobacman and Cammarato, 2021). Fig. 8 indicates these cooperative interactions among the filaments and substantial isoform differences. In the case of ssTnI compared with cTnI, these differences occur in the critical inhibitory peptide (Ip), switch peptide (Sw), and mobile C-terminal domains. cTnT-R92Q is in a critical helical region (cTnT-87-150) that provides an anchor to the thin filament essential in the transduction of the Ca2+-binding signal and the interaction between contiguous peptides in the Tm strand (Tobacman and Cammarato, 2021).

Mechanisms of regulation of cardiac myofilament activation modified by myosin activators and inhibitors. As shown by the right side of the figure, the reaction of cross-bridges with thin filaments promotes ATP hydrolysis, powering shortening and tension generation. At the level of the thin filaments, Ca-binding to cTnC triggers an interaction of a Sw of the inhibitory protein (TnI) to switch on the thin filament and release the inhibition imposed by a TnI-inhibitory peptide (Ip), the mobile domain of TnI, and TnT interactions with actin-Tm interface. The result is a movement of Tm permitting the actin-cross-bridge reaction. The extent of myofilament activation is governed by the number of cross-bridges reacting with the thin filament, which is determined by the proportion of cTnC sites with bound Ca2+ related to the amounts of Ca2+ released and to the population of cross-bridges existing in an SRX and a DRX. As shown by the green arrow extending along the myofilaments, the extent of activation is also determined by the cooperative spread of activation from an active regulatory unit to near neighbors among the regulatory units. Cross-bridge interactions with thin filament promote this spread, as do modifications in energies of interaction among neighboring thin filament proteins, especially interactions at the ends of contiguous Tm strands. Critical regions of TnI differ between cTnI and ssTnI. The N-terminal extension is specific to cTnI. In homologous regions the cTnI Ip T143 is P111 in ssTnI, in the cTnI Sw Q155, A162 and E164 are R125, H132, V134 in ssTnI, and in the cTnI mobile domain cTnI H172 and K207 are N141 and M175 in ssTnI. Data from our studies indicate that myosin activators and inhibitors are affected by this difference, which affect activation and inhibition of tension. See text for further discussion.
The mechanisms currently proposed for the actions of OM stress its effects on myosin kinetics, the dwell time of the force generating cross-bridges on the thin filament, or the population of cross-bridges in SRX and DRX states. There has been little consideration of the potential that these effects are influenced by the thin filaments, their isoforms, or proteins associated with the thick filaments, such as cardiac myosin binding protein C. Similarly, with Mava the proposed mechanism focuses on shifts in the population of cross-bridges to the SRX state (Anderson et al., 2018).
Molecular Mechanisms of OM in Developing Cardiac Sarcomeres
Effects of OM in our studies included increases in submaximal tension with modifications of cooperative activation and Ca2+-sensitivity. The effect of OM to increase submaximal tension developed by cTnI myofilaments compared with ssTnI myofilaments stresses how a change in cooperativity has impactful effects on tension developed in addition to modifications in Ca2+-sensitivity. In agreement with our studies with cardiac fibers, previous studies on slow skeletal fibers reported OM substantially increasing Ca2+- sensitivity, with a large effect at submaximal Ca2+ concentrations and a decrease in nH (Nagy et al., 2015; Lindqvist et al., 2019; Governali et al., 2020). As illustrated in Fig. 8, OM is likely to modify cooperative activation by a mechanism involving an interaction of force generating cross-bridges with thin filaments inducing movements in Tm or altered actin-actin interactions modifying the spread of activation from an active regulatory unit to a near-neighbor regulatory unit. The influence of effects of OM on the interaction between myosin S-2 and cardiac myosin binding protein C may also be a factor (Mamidi et al., 2015). Although isoform shifts were not studied, there are reports supporting our conclusion that thin filament activation is at play in the effects of OM (Kampourakis et al., 2018; Woody et al., 2018). Woody et al. (2018) reported that the increase in force induced by OM is associated with a suppression of the working stroke of cardiac myosin together with thin filament activation. Kampourakis et al. (2018) reported that the modification of cross-bridge state induced by OM in adult skinned fibers also increased Ca2+-sensitivity. Mamidi et al. (2015) reported the effects of OM to alter function in skinned fibers lacking MyBP-C in a way suggesting that it may be useful in hypercontractile states in HCM models with deletion mutations in MyBP-C. In this case the actions of OM are the opposite of its proposed effects to enhance contractility.
Molecular Mechanisms of Mava in Inhibition of Developing Cardiac Sarcomeres
Data presented here are the first to report the effects of Mava on Ca2+-activation of skinned ventricular fibers expressing either cTnI or ssTnI with and without expression of an HCM-linked mutant, cTnT-R92Q. Inhibition of maximum and submaximum tension values was the same in adult myofilaments expressing either cTnI or ssTnI in place of cTnI. However, the influence of the thin filament state on the effect of Mava was indicated by the demonstration that in the presence of Mava, nH values were different in NTG fibers than in TG-ssTnI fibers. The use of probes as reporters of Ca2+ localized at the thin filament indicate an effect of Mava on thin filament activation that may be different in cTnI-sarcomeres versus second TnI-sarcomeres. These studies report little change in the peak values of Ca2+-troponin binding induced by Mava, but a considerable effect on the rate of binding and release from troponin (Sparrow et al., 2019). This interpretation is tempered by the use of unloaded myocytes. Sparrow et al. (2020) reported no effect of Mava on ATPase rate in control unloaded reconstituted myofilament preparations and a very small effect in preparations with thin-filament HCM-linked mutations. A weakness of these studies is the use of unloaded reconstituted preparations lacking critical proteins in the myofilament lattice.
Therapy Development and Effects of OM and Mava in Immature Myofilaments
Importantly, our study expands the understanding of how OM and Mava affect sarcomere tension, which supports the need for rethinking the mechanisms of their actions and those of new sarcomere activators and inhibitors likely to be developed. An encouraging aspect of the application of data presented here for future drug development is the success of safety trials with OM and Mava. As reported by Teerlink et al. (2020; 2021), there were minimal side effects in the OM clinical trials. Similarly, Olivotto et al. (2020) reported minimal side effects of Mava in the EXPLORER-HCM clinical trial. A recent study (Barrick et al., 2021), relevant to those reported here, emphasizes the significance of understanding detailed mechanisms of action of sarcomere activators and inhibitors in precision medicine applied in pediatric cardiology. They investigated one of several mutants (cTnT-R134G) linked to pediatric dilated cardiomyopathy (DCM) progressing to pediatric heart failure (Hershberger et al., 2009). In myofilaments controlled by cTnT-R134G, Ca2+-cTnC was unable to switch on the myofilaments, and cross-bridge interactions could not activate the thin filament. As pointed out by Barrick et al. (2021), in this cardiac disorder, the use of OM to increase contractility, which we show can activate the thin filament in immature myofilaments, would be preferred over activators (e.g., levosimendan) promoting transduction of the cTnC Ca2+-binding signal (Klein et al., 2018).
In summary, despite different mechanisms of action to activate and inhibit sarcomere tension, effects of OM and Mava are similar in immature and mature myofilaments. However, the mechanisms in mature and immature sarcomeres involve not only induction of DRX and SRX states, but also modifications in the mechanism related to cooperative activation of the myofilaments. The results are also related to screening for sarcomere activators and inhibitors using cardiac myocytes derived from human induced pluripotent stem cell (hiPSCs), which are generally immature and express ssTnI. A relevant finding reported in our previous studies is the demonstration that OM induced activation of sarcomeres from hearts of a DCM model (Tm-E54K) (Utter et al., 2015), in human induced pluripotent stem cell-derived cardiomyocytes (hiPSCs-CMs) expressing ssTnI, and DCM mutant cTnT R173W (Broughton et al., 2016). More recently, (Rhoden et al., 2022) also reported effects of OM on engineered human cardiac preparations likely to contain ssTnI.
Authorship Contributions
Participated in research design: Halas, Langa, Warren, Solaro.
Conducted experiments: Halas, Warren.
Performed data analysis: Halas, Langa, Warren, Solaro.
Wrote or contributed to the writing of the manuscript: Halas, Langa, Warren, Goldspink, Wolska, Solaro.
Footnotes
- Received September 28, 2021.
- Accepted February 16, 2022.
↵1 M.H. and P.L. contributed equally to this work.
This work was supported by National Institutes of Health National Heart Lung and Blood Institute [Grant R01-HL128468] to J.S. and B.M.W. and [Grant P01-HL062426] to R.J.S., C.M.W., B.M.W., and P.H.G.; and the American Heart Association [Grant 834691] to P.L.
Dr. Solaro is a member of the Scientific Advisory Board of Cytokinetics and a consultant to MyoKardia BMS and Pfizer, Inc.
↵
 This article has supplemental material available at molpharm.aspetjournals.org.
This article has supplemental material available at molpharm.aspetjournals.org.

Abbreviations
- cTnC
- cardiac troponin C
- cTnI
- cardiac troponin I
- cTnT
- cardiac troponin T
- DCM
- dilated cardiomyopathy
- DRX
- disordered relaxed state
- HCM
- hypertrophic cardiomyopathy
- HR
- high-relaxing buffered solution
- Ip
- inhibitory peptide
- Mava
- Mavacamten
- nH
- Hill slope
- NTG
- nontransgenic
- OM
- omecamtiv mecarbil
- R92Q
- Arg92Gln mutation in TnT
- SRX
- super relaxed state
- ssTnI
- slow skeletal troponin I
- Sw
- switch peptide
- TG
- transgenic
- Tm
- tropomyosin
- TnI
- troponin I
- TnT
- troponin T
- Copyright © 2022 by The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}