


Article Figures & Data
Figures
- Fig. 1.
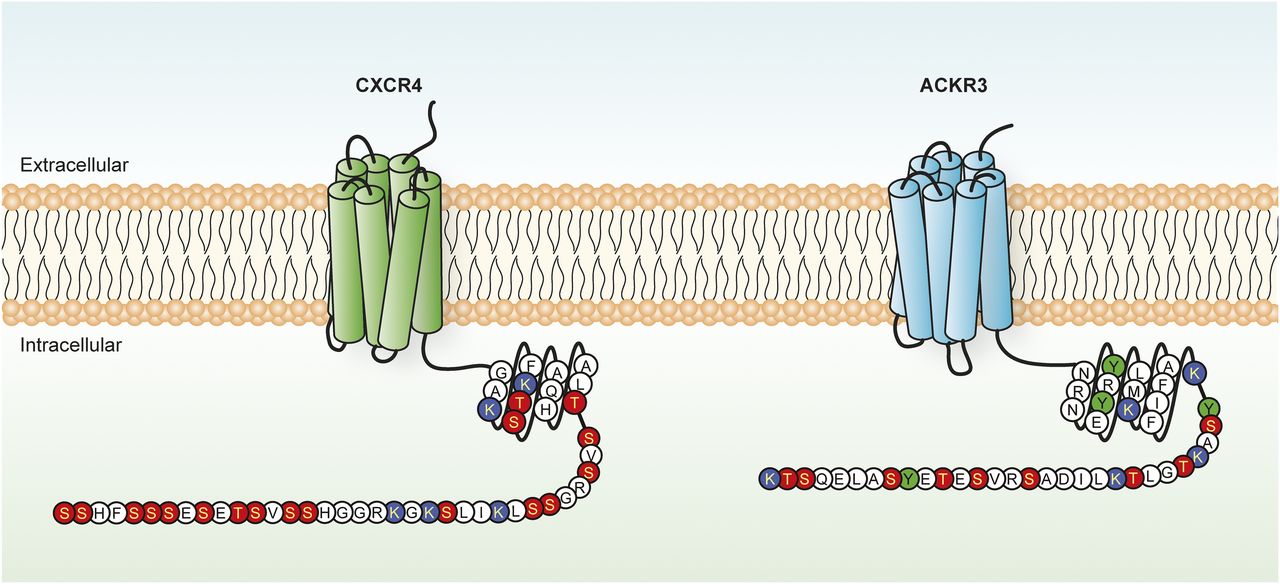
CXCR4 and ACKR3 residues potentially subjected to post-translational modifications. Schematic representation of the C-terminal tail of CXCR4 and ACKR3, in which serine/threonine (red), tyrosine (green), and lysine (blue) residues potentially subjected to post-translational modifications are highlighted.
- Fig. 2.
CXCR4 C-terminus phosphosites. Schematic representation of the C-terminal tail of CXCR4, in which serine residues known to be phosphorylated are highlighted in light blue. The kinases or the extracellular stimuli responsible for the phosphorylation are also specified. Hrg, heregulin.

Tables
- TABLE 1
Principal methods used to identify GPCR-interacting proteins and phosphorylated residues.
GPCR-interacting proteins (1–4) and phosphorylated residues (5).
Classification Method Screening Advantages Disadvantages 1. Genetic Y2H Highly suitable Easy to perform. Inexpensive. Loss of spatial-temporal information. Membrane-anchored proteins cannot be investigated. Performed in yeast. MYTH Highly suitable Easy to perform. Membrane-anchored proteins can be investigated. Loss of spatial-temporal information. Soluble proteins cannot be investigated. Performed in yeast. MaMTH Highly suitable Easy to perform. Membrane anchored proteins can be investigated. Performed in mammalian cells. Loss of spatial-temporal information. Soluble proteins cannot be investigated. KISS Possible Sensitive enough for studying interaction dynamic. Loss of spatial-temporal information. Both membrane and cytosolic proteins can be investigated. Proteins involved in the STAT3 cascade cannot be investigated. 2. Biophysical BRET/FRET Not suitable Precise spatial-temporal information. High sensitivity. Possibility to study interactions in living cells. Generation of fusion proteins. Relies on the proximity and relative orientation between donor and acceptor. Fluorescent lifetime microscopy Suitable More accurate than intensity-based FRET. Data analysis more laborious than intensity-based FRET. 3. Biochemical PLA Not suitable Precise spatial information (single-molecule resolution). Possibility to perform in ex-vivo models. Relies on antibodies. High cost. Not easy to scale up in large studies. BioID Suitable Precise spatial information. Not well suited for studying interaction dynamic (fluorescent signal is delayed). Several interactions in parallel. Possibility to perform in living cells. NanoBit Suitable Precise spatial information. Several interactions in parallel. Possibility to perform in living cells. 4. Proteomic Co-IP Highly suitable Purification of protein complexes in living cells and tissues. Rely on antibodies. Loss of spatial-temporal information. Lysis conditions might influence results. Pull-down Highly suitable Can prove direct interaction. Loss of spatial-temporal information. In vitro binding assays. Fusion of the receptor on the beads might alter receptor conformation. BioID Highly suitable Can detect weak and transient interactions in living cells. Fusion of the biotin to the receptor might alter its targeting or functions. 5. Phosphorylation [32P] Suitable Very sensitive. Radioactive method. Cannot give information on the number of phosphorylated residues nor their position. LC-MS Highly suitable Can pinpoint phosphorylated residues. Can yield false negatives. Not quantitative unless combined with very expensive isotope tags. Mutagenesis Suitable Cheap and easy. Indirect method. Based on functional data in living cells. Can pinpoint phosphorylated residues. Mutagenesis of the C-terminus can impair expression and/or localization of the receptor. Labor-intensive in case of multiple phosphosites. Not quantitative. Phospho-antibodies Suitable Direct and indirect. Can be used in any cell line. Semiquantitative and qualitative. Time consuming and expensive for the generation of the antibodies. Useless with low affinity antibodies. Cannot give information on contiguous phosphorylated residues. BiFC, bimolecular fluorescent complementation assay; BioID, proximity-dependent biotin identification; KISS, kinase substrate sensor; MaMTH, mammalian membrane two-hybrid assay; MYTH, membrane yeast two-hybrid assay; PLA, proximity ligation assay; Y2H, yeast two-hybrid assay.
Protein Method of Identification Cellular Context Direct Constitutive/Induced Site of Interaction Role Reference CXCR4-interacting proteins Filamin A Pull-down HEK293 cells Yes Constitutive and CXCL12-induced The ROCK inhibitor Y27632 reverses CXCL12-induced increased interaction C-terminal tail and third loop of CXCR4 Stabilize CXCR4 at the surface Gómez-Moutón et al. (2015) Co-IP Recombinant protein AIP4 Pull Down HEK293 cells Yes Constitutive and CXCL12-induced CXCR4 C-tail serines and WW domains of AIP4. Serine 324 and 325 when phosphorylated increase interaction Increase CXCR4 degradation Bhandari et al. (2009) Co-IP FRET RTN3 Y2H HEK293 cells NA Constitutive, induction not tested Carboxyl terminal of RTN3 Increase cytoplasmic localization of CXCR4 Li et al. (2016) Co-IP CD74 Co-IP HEK293 and MonoMac6 cells NA Constitutive, induction not tested NA Phosphorylation of AKT Schwartz et al. (2009) Colocalization TLR2 FRET Human monocyte and HEK293 cells NA Induced by Pg-fimbria NA CXCR4 inhibits TLR2-induced NF- κB activation. In addition, CXCR4 found to be receptor of the pattern-recognition receptor complex Hajishengallis et al. (2008), Triantafilou et al. (2008) Co-IP NMMHC Pull-down Jurkat T and Peer T cells lymphocytes NA Constitutive and not induced by CXCL12 CXCR4 C-terminus Lymphocytes migration Rey et al. (2002) Co-IP Colocalization Drebrin Pull Down J77 T, YES Constitutive and induced by superantigen E, which also relocalizes the interaction to the leading edge of migrating lymphocytes Drebrin Drebrin affects key physiologic processes during antigen presentation in HIV entry Pérez-Martínez et al. (2010), Gordón-Alonso et al. (2013) Co-IP HEK293T and N-terminus positively regulates interaction whereas the C-terminus seems to negatively regulate it FRET HIV-infected T cells CD164 Co-IP Jurkat and NA Only induced when CXCL12 is presented on fibronectin NA CD164 participates to the CXCL12 mediated AKT and PKC-ζ phosphorylation Forde et al. (2007), Huang et al. (2013) Colocalization Ovarian surface epithelial cells mDIA2 Co-IP MDA-MB-231 cells NA Constitutive (very weak) and CXCL12 induced NA Cytoskeletal rearrangement necessary for nonapoptotic blebbing Wyse et al. (2017) Colocalization ATP13A2 MYTH Yeast YES Constitutive NA NA Usenovic et al. (2012) PI3Kγ Co-IP Human myeloid cells NA Only CXCL12 induced NA Integrin activation and chemotaxis Schmid et al. (2011) PECAM-1 PLA Human coronary artery endothelial cells NO Constitutive Induction not studied NA CXCR4 part of a junctional mechano-sensitive complex dela Paz et al. (2014) MYBL2 2HY Yeast Yes NA NA NA Wang et al. (2011) KCNK1 Co-IP HeLa cells NA NA NA NA Hein et al. (2015) TMEM63B Co-IP HeLa cells NA NA NA NA Hein et al. (2015) GOLT1B Co-IP HeLa cells NA NA NA NA Hein et al. (2015) eIFB2 Co-IP Pre-B NALM6 cells NA Constitutive and negatively regulated by CXCL12 NA NA Palmesino et al. (2016) Colocalization CDC73 Co-IP Pre-B NALM6 cells NA Constitutive Induction not studied NA NA Palmesino et al. (2016) CTR9 Co-IP Pre-B NALM6 cells NA Constitutive Induction not studied NA NA Palmesino et al. (2016) PAF1 Co-IP Pre-B NALM6 cells NA Constitutive Induction not studied NA NA Palmesino et al. (2016) NPM Co-IP Pre-B NALM6 cells NA Constitutive Induction not studied NA NA Palmesino et al. (2016) CD11B Co-IP Pre-B NALM6 cells NA Constitutive Induction not studied NA NA Palmesino et al. (2016) PTPRS Co-IP Pre-B NALM6 cells NA Constitutive Induction not studied NA NA Palmesino et al. (2016) LGALS8 Co-IP Pre-B NALM6 cells NA Constitutive Induction not studied NA NA Palmesino et al. (2016) ACKR3-interacting proteins EGFR PLA MCF7 cells, breast cancer tissues, CaP cells Mediated by β-arrestin2 Interaction constitutive and induced by the epidermal growth factor. NA ACKR3 mediates phosphorylation of EGFR at tyrosine1110 after EGF-stimulation and phosphorylation of ERK1/2 with consequences on tumor proliferation. Singh and Lokeshwar (2011), Salazar et al. (2014), Kallifatidis et al. (2016) Colocalization Co-IP CD74 Co-IP NIH/3T3 cells and human B cells NA Constitutive, induction not investigated. NA ACKR3 is involved in MIF-mediated ERK-1/2 and ζ-chain-associated protein kinase activation in addition to MIF-mediated chemotaxis. Alampour-Rajabi et al. (2015) Colocalization PLA PECAM-1 PLA HCAECs cells NA Constitutive NA NA dela Paz et al. (2014) ATP13A2 MYTH Yeast Yes NA NA NA Usenovic et al. (2012) GJB2 Co-IP HeLa cells NA NA NA NA Hein et al. (2015) MRPL4 Co-IP HeLa cells NA NA NA NA Hein et al. (2015) ATP5H Co-IP HeLa cells NA NA NA NA Hein et al. (2015) ATP5B Co-IP HeLa cells NA NA NA NA Hein et al. (2015) ATP5A1 Co-IP HeLa cells NA NA NA NA Hein et al. (2015) ATP5O Co-IP HeLa cells NA NA NA NA Hein et al. (2015) ACKR3 Co-IP HeLa cells NA NA NA NA Hein et al. (2015) ESPL1 Co-IP HeLa cells NA NA NA NA Hein et al. (2015) HECTD2 Co-IP HeLa cells NA NA NA NA Hein et al. (2015) UBR7 Co-IP HeLa cells NA NA NA NA Hein et al. (2015) ATP13A2, cation-transporting ATPase 13A2; ATP5H/ATP5B/ATP5A1/ATP5O, ATP synthase subunit d/beta/alpha/O, mitochondrial; CAP, human prostate cancer cells; CD11B, cyclin-dependent kinase 11B; CD164, endolyn; CDC73, parafibromin; CTR9, SH2 domain binding protein; eIF2B, eukaryotic translation initiation factor 2B; EGFR, epidermal growth factor receptor; ESPL1, HCAEC, Human Coronary Artery Endothelial Cells; HECTD2, probable E3 ubiquitin-protein ligase; GJB2, gap junction β-2 protein; GOLT1B, vesicle transport protein GOT1B; KCNK1, potassium channel subfamily K member 1; LGALS8, galectin; mDIA2, diaphanous-related formin-2; MYBL2, myb-related protein B; MRPL4, 54S ribosomal protein L4, mitochondrial; MYTH, membrane yeast two-hybrid assay; NA, not applicable; NF, nuclear factor; NMMHC, motor protein nonmuscle myosin H chain; NPM, nucleophosmin; PAF1, hypothetical protein PD2; PECAM-1, platelet endothelial cell adhesion molecule; PI3Kγ, PI3-kinase isoform p110γ; PLA, proximity ligation assay; PTPRS, receptor-type tyrosine-protein phosphatase S; RTN3, reticulon3; TLR2, Toll-like receptor 2; TMEM63B, CSC1-like protein 2; UBR7, putative E3 ubiquitin-protein ligase; Y2H, yeast two-hybrid assay.




{kind=link}
{kind=link}
{kind=link}